

Interaction between IGF-IR and ER induced by E2 and IGF-I
- PMID: 23704881
- PMCID: PMC3660452
- DOI: 10.1371/journal.pone.0062642
Interaction between IGF-IR and ER induced by E2 and IGF-I
Abstract
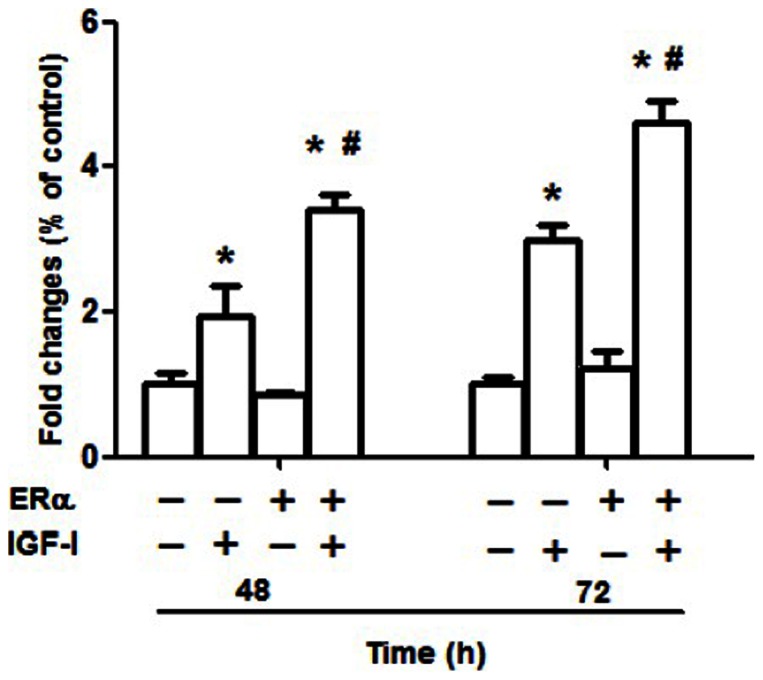
Estrogen receptor (ER) is a nuclear receptor and the insulin-like growth factor-I (IGF-I) receptor (IGF-IR) is a transmembrane tyrosine kinase receptor. Estrogen and IGF-I are known to have synergistic effects on the growth of breast cancer cells. Recently, non-nuclear effects of ER have been under investigation. To study the mechanism involved in this process, we have used MCF-7 breast cancer cell lines transfected with IGF-IR anti-sense cDNA (SX13, MCF-7(SX13)) that resulted in 50% reduction of IGF-IR. In MCF-7 cells, estradiol (E2) and IGF-I induced the rapid association of ER to IGF-IR, however, the interaction was abrogated in MCF-7(SX13) cells. In addition, NWTB3 cells (NIH3T3 cells overexpressing IGF-IR) were transiently transfected with ERα, the ER-IGF-IR interaction was induced by both E2 and IGF-I. Moreover, ERα regulated the IGF-I signaling pathways through phosphorylation of ERK1/2 and Akt and the interaction of ER-IGF-IR potentiated the cell growth. Finally, E2 and IGF-I stimulated translocation of ER from the nucleus to the cytoplasm. Taken together, these findings reveal that the interaction of the ER and IGF-IR is important for the non-genomic effects of ER.
Conflict of interest statement
Figures
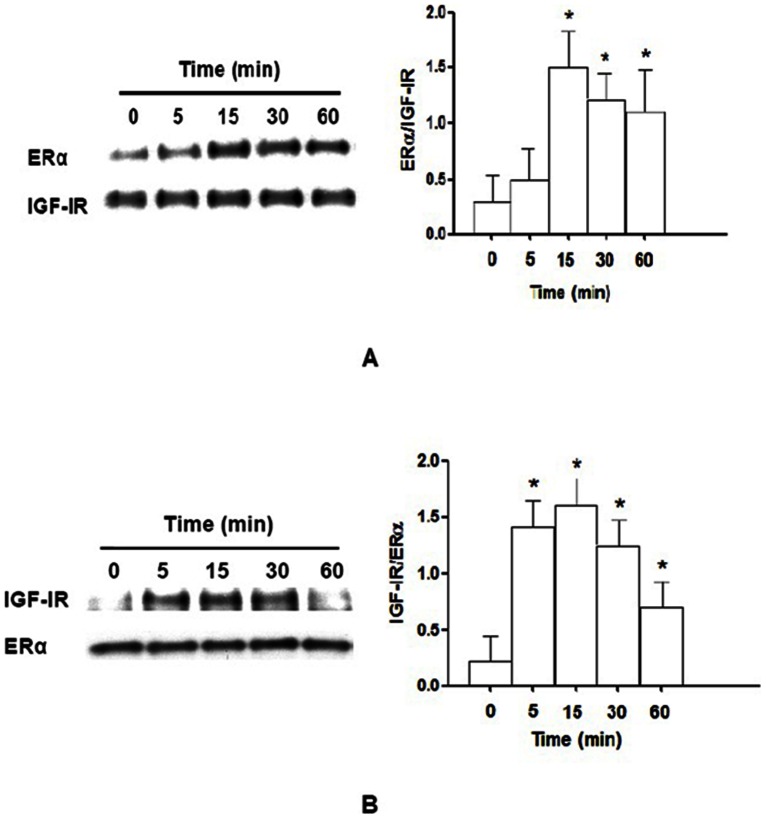
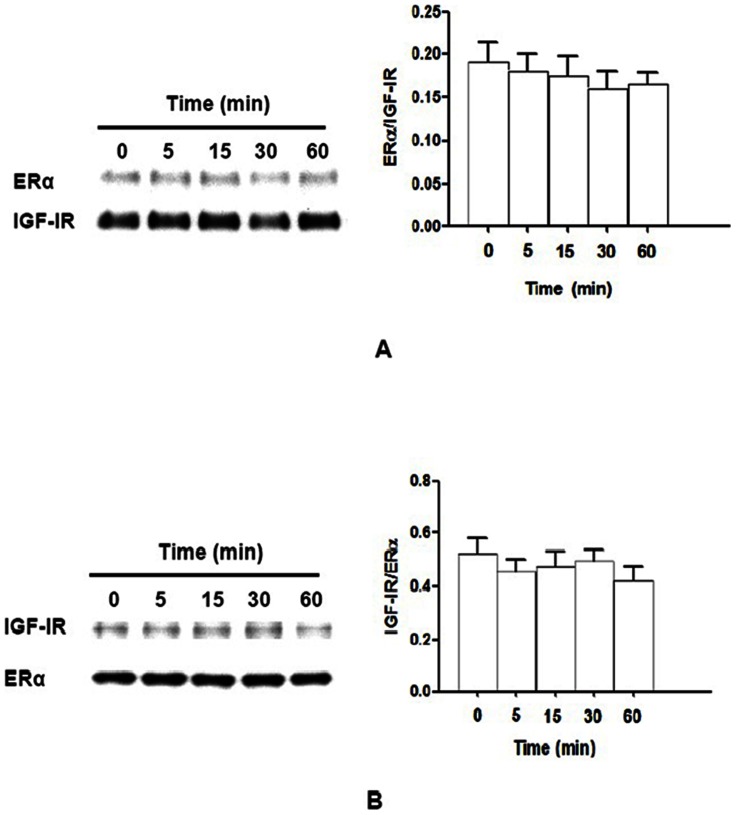
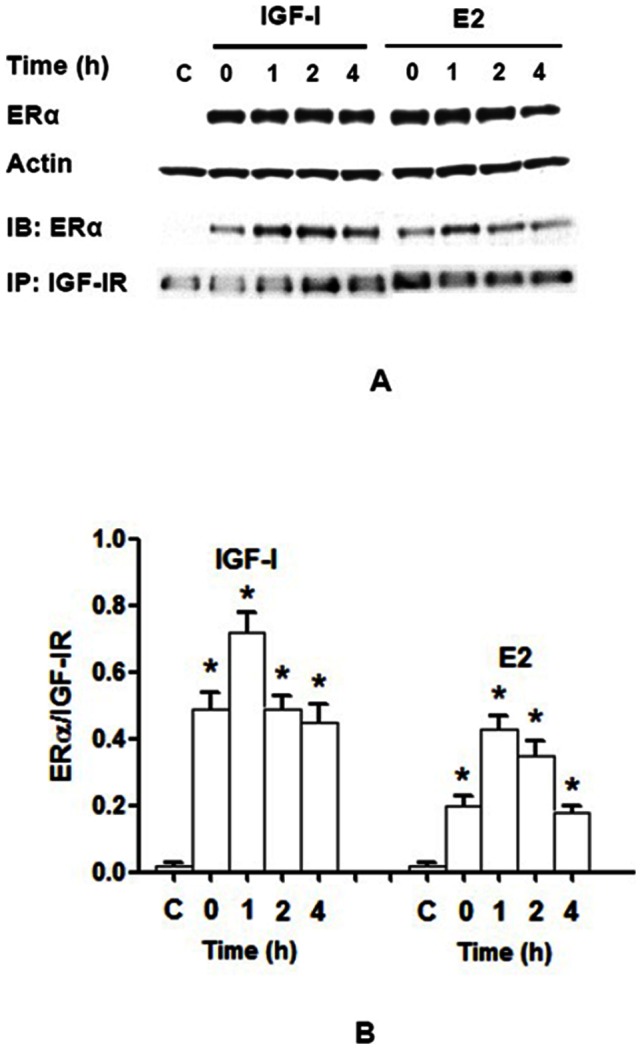
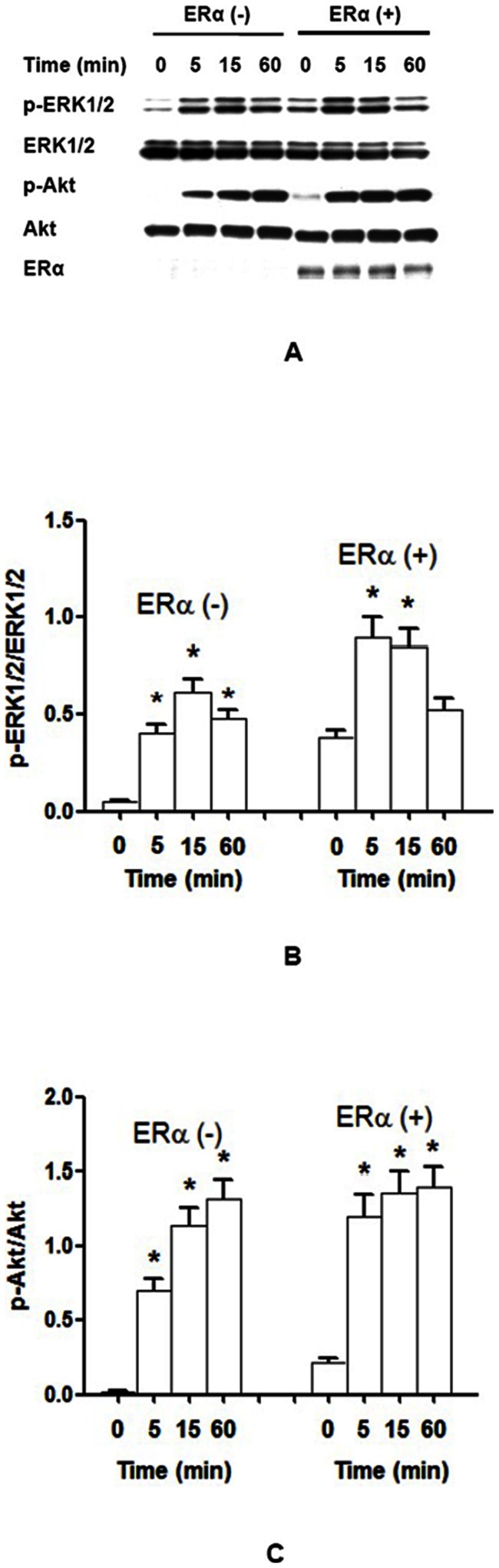

References
-
- Kawabe Y, Kuwayama H, Morio T, Urushihara H, Tanaka Y (2002) A putative serpentine receptor gene tasA required for normal morphogenesis of primary stalk and branch structure in Polysphondylium pallidum. Gene 285: 291–9. - PubMed
-
- Stenoien DL, Mancini MG, Patel K, Allegretto EA, Smith CL, et al. (2000) Subnuclear trafficking of estrogen receptor-alpha and steroid receptor coactivator-1. Mol Endocrinol 14: 518–34. - PubMed
-
- Bartella V, De Marco P, Malaguarnera R, Belfiore A, Maggiolini M (2012) New advances on the functional cross-talk between insulin-like growth factor-I and estrogen signaling in cancer. Cell Signal 24: 1515–21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
