

Tetraspanins CD81 and CD82 facilitate α4β1-mediated adhesion of human erythroblasts to vascular cell adhesion molecule-1
- PMID: 23704882
- PMCID: PMC3660455
- DOI: 10.1371/journal.pone.0062654
Tetraspanins CD81 and CD82 facilitate α4β1-mediated adhesion of human erythroblasts to vascular cell adhesion molecule-1
Abstract
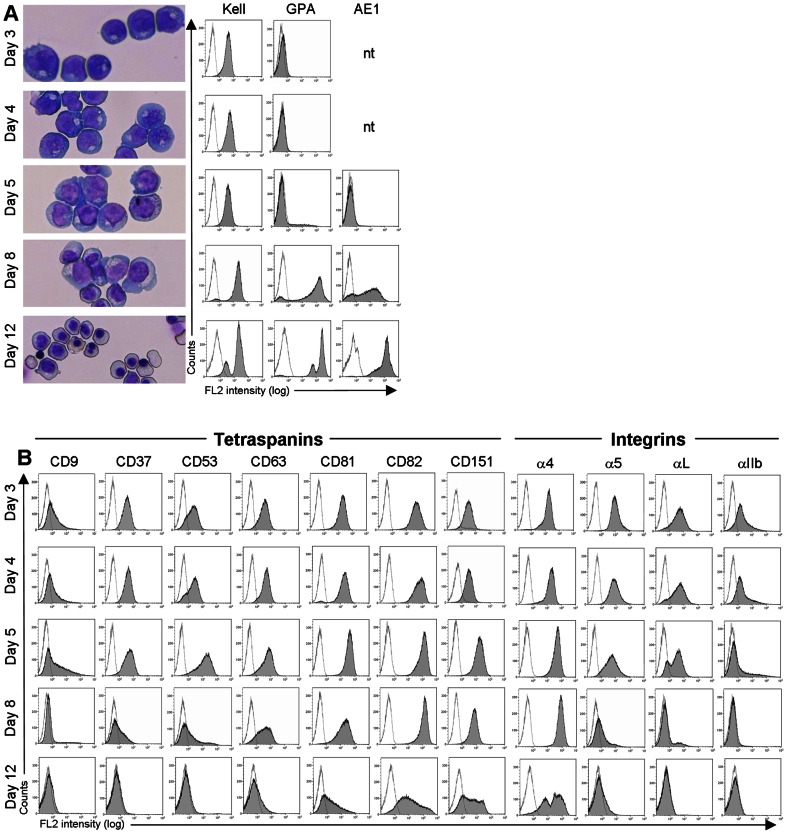
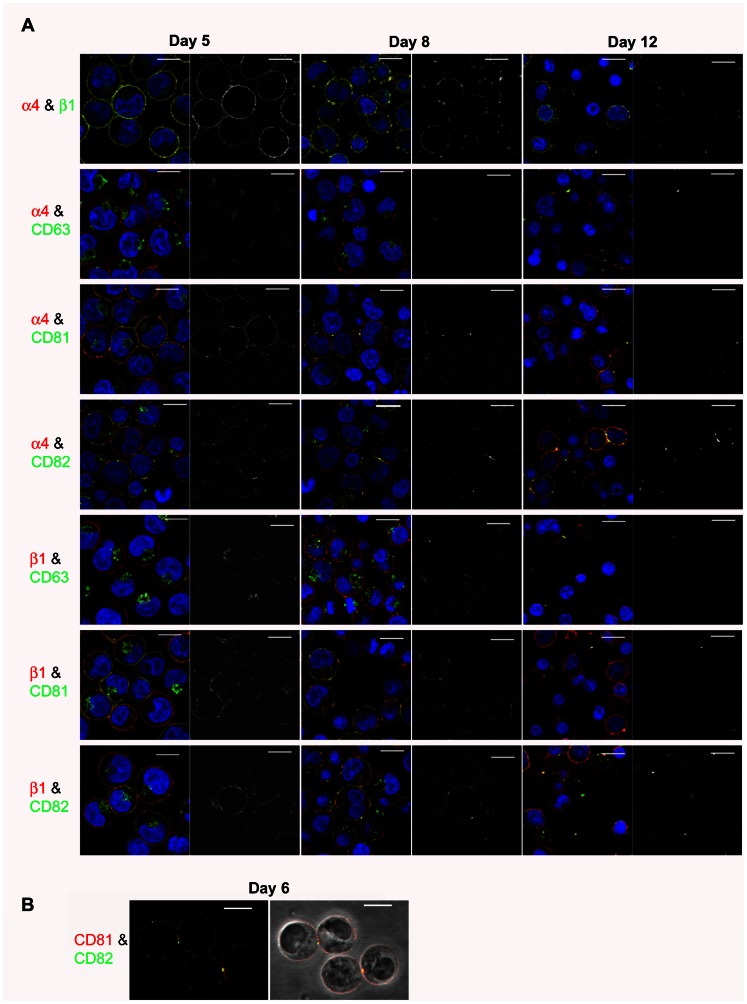
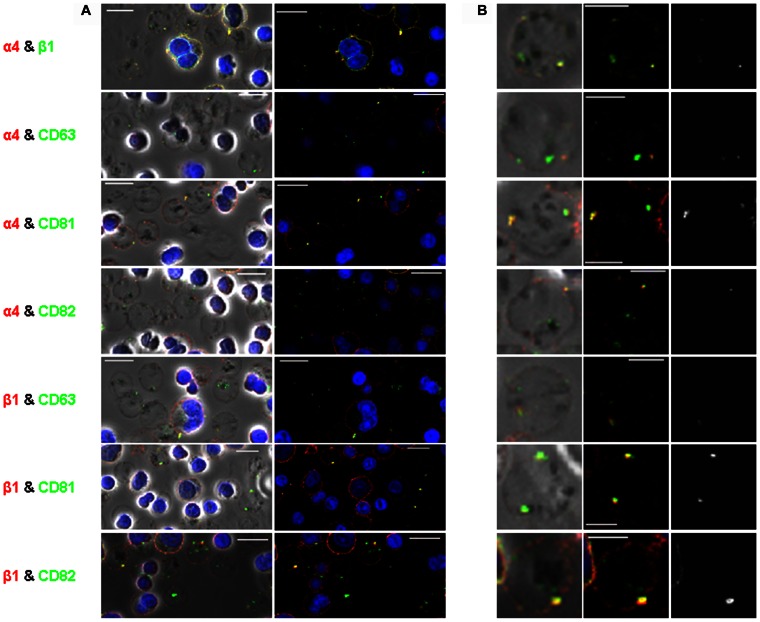
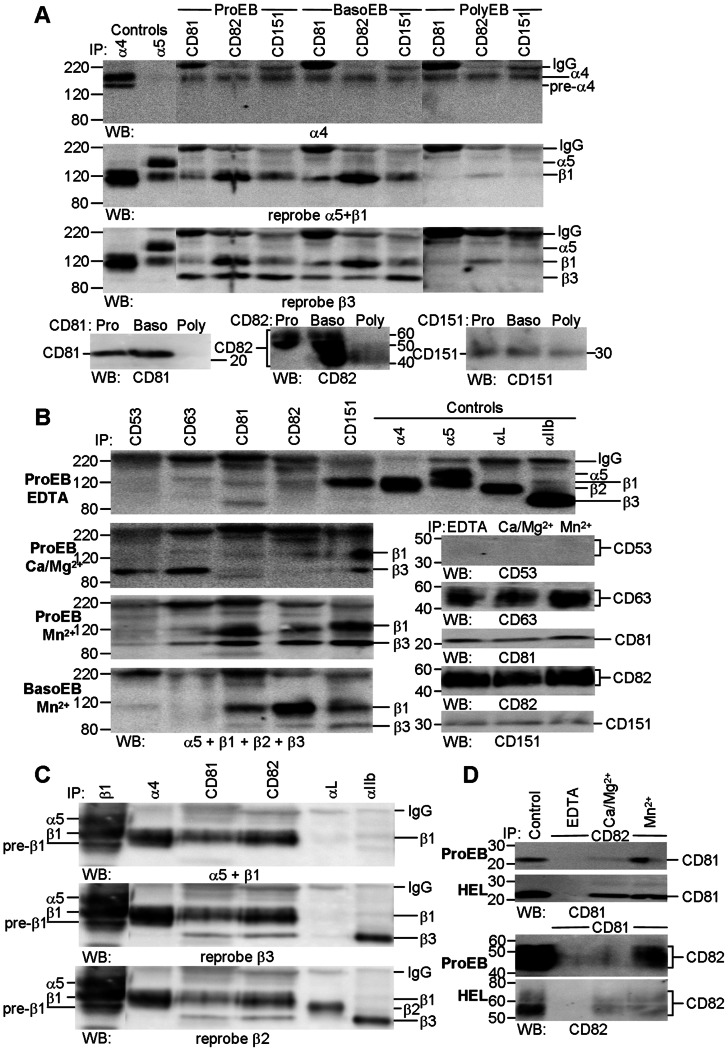
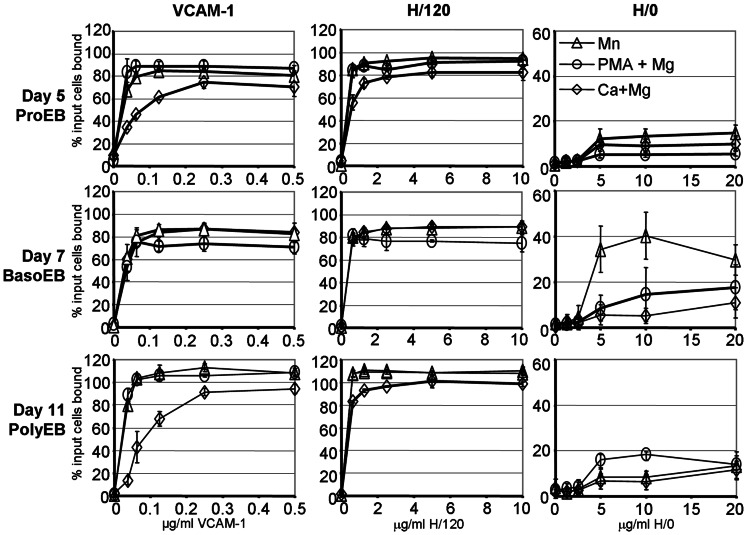
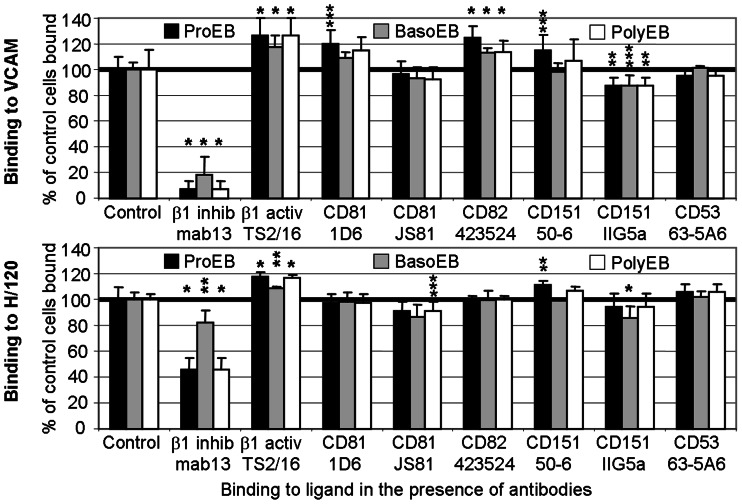
The proliferation and terminal differentiation of erythroid progenitors occurs in human bone marrow within erythroblastic islands, specialised structures consisting of a central macrophage surrounded by developing erythroid cells. Many cell-cell and cell-matrix adhesive interactions maintain and regulate the co-ordinated daily production of reticulocytes. Erythroid cells express only one integrin, α4β1, throughout differentiation, and its interactions with both macrophage Vascular Cell Adhesion Molecule-1 and with extracellular matrix fibronectin are critical for erythropoiesis. We observed that proerythroblasts expressed a broad tetraspanin phenotype, and investigated whether any tetraspanin could modulate integrin function. A specific association between α4β1 and CD81, CD82 and CD151 was demonstrated by confocal microscopy and co-immune precipitation. We observed that antibodies to CD81 and CD82 augmented adhesion of proerythroblasts to Vascular Cell Adhesion Molecule-1 but not to the fibronectin spliceoforms FnIII12-IIICS-15 and FnIII12-15. In contrast, different anti-CD151 antibodies augmented or inhibited adhesion of proerythroblasts to Vascular Cell Adhesion Molecule-1 and the fibronectin spliceoform FnIII12-IIICS-15 but not to FnIII12-15. These results strongly suggest that tetraspanins have a functional role in terminal erythropoiesis by modulating interactions of erythroblast α4β1 with both macrophages and extracellular matrix.
Conflict of interest statement
Figures
References
-
- Bessis M (1958) [Erythroblastic island, functional unity of bone marrow]. Rev Hematol 13: 8–11. - PubMed
-
- Mohandas N, Prenant M (1978) Three-dimensional model of bone marrow. Blood 51: 633–643. - PubMed
-
- Morris L, Crocker PR, Fraser I, Hill M, Gordon S (1991) Expression of a divalent cation-dependent erythroblast adhesion receptor by stromal macrophages from murine bone marrow. J Cell Sci 99: 141–147. - PubMed
-
- Hanspal M, Hanspal JS (1994) The association of erythroblasts with macrophages promotes erythroid proliferation and maturation: a 30-kD heparin-binding protein is involved in this contact. Blood 84: 3494–3504. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
