

The weak shall inherit: bacteriocin-mediated interactions in bacterial populations
- PMID: 23704942
- PMCID: PMC3660564
- DOI: 10.1371/journal.pone.0063837
The weak shall inherit: bacteriocin-mediated interactions in bacterial populations
Abstract
Background: Evolutionary arms race plays a major role in shaping biological diversity. In microbial systems, competition often involves chemical warfare and the production of bacteriocins, narrow-spectrum toxins aimed at killing closely related strains by forming pores in their target's membrane or by degrading the target's RNA or DNA. Although many empirical and theoretical studies describe competitive exclusion of bacteriocin-sensitive strains by producers of bacteriocins, the dynamics among producers are largely unknown.
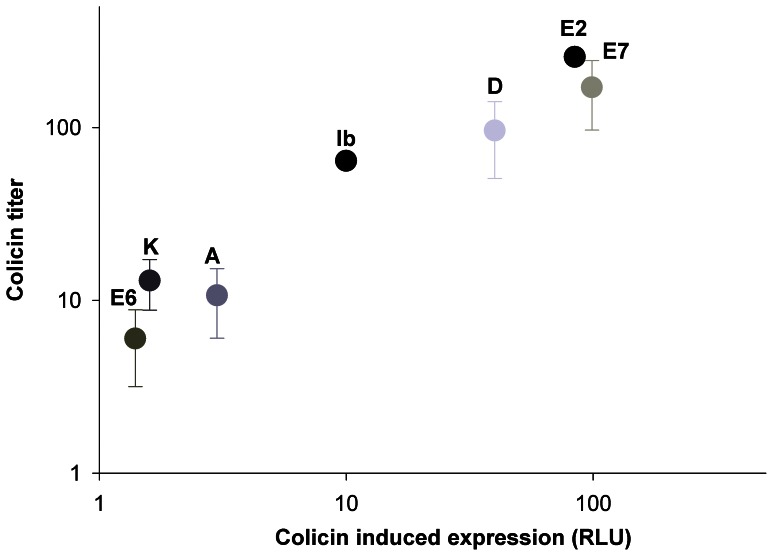
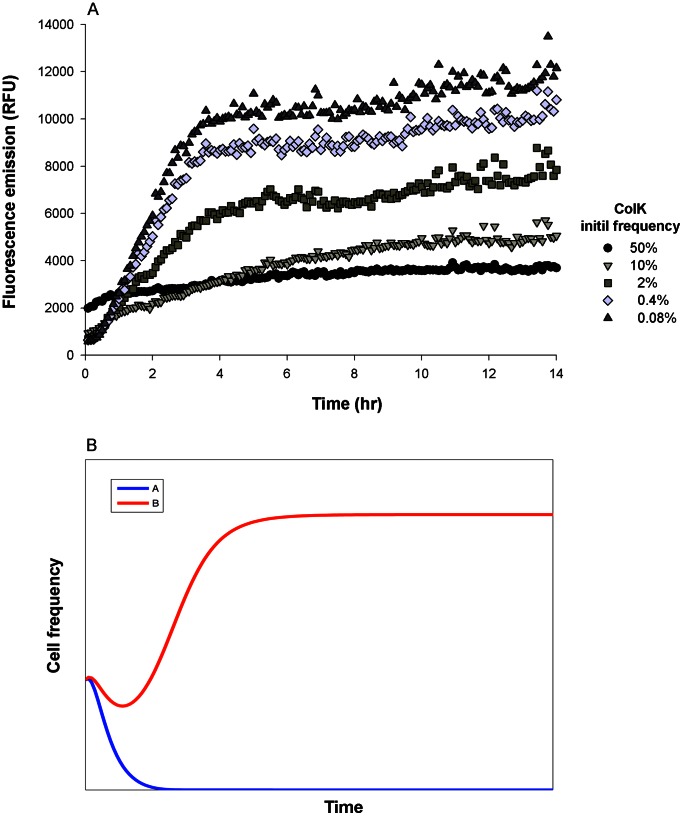
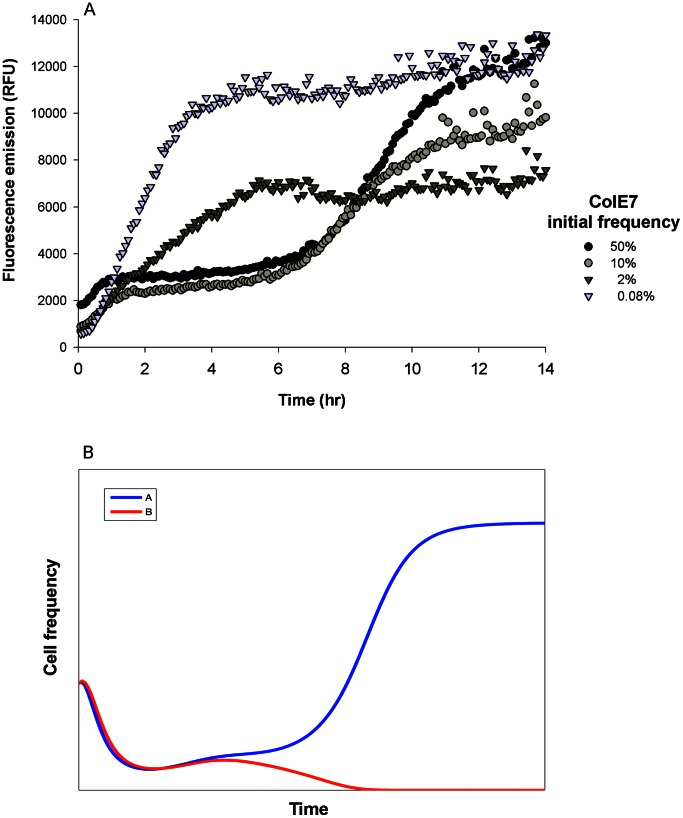
Methodology/principal findings: We used a reporter-gene assay to show that the bacterial response to bacteriocins' treatment mirrors the inflicted damage Potent bacteriocins are lethal to competing strains, but at sublethal doses can serve as strong inducing agents, enhancing their antagonists' bacteriocin production. In contrast, weaker bacteriocins are less toxic to their competitors and trigger mild bacteriocin expression. We used empirical and numerical models to explore the role of cross-induction in the arms race between bacteriocin-producing strains. We found that in well-mixed, unstructured environments where interactions are global, producers of weak bacteriocins are selectively advantageous and outcompete producers of potent bacteriocins. However, in spatially structured environments, where interactions are local, each producer occupies its own territory, and competition takes place only in "no man's lands" between territories, resulting in much slower dynamics.
Conclusion/significance: The models we present imply that producers of potent bacteriocins that trigger a strong response in neighboring bacteriocinogenic strains are doomed, while producers of weak bacteriocins that trigger a mild response in bacteriocinogenic strains flourish. This counter-intuitive outcome might explain the preponderance of weak bacteriocin producers in nature. However, the described scenario is prolonged in spatially structured environments thus promoting coexistence, allowing migration and evolution, and maintaining bacterial diversity.
Conflict of interest statement
Figures
 , and the initial relative frequency varied
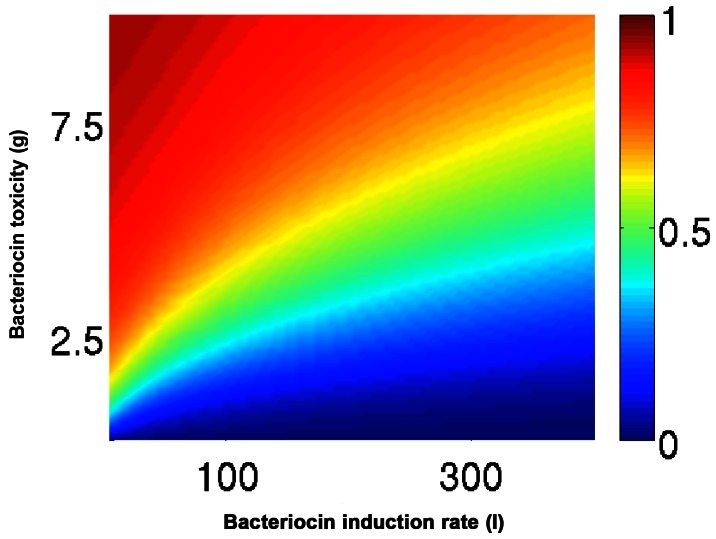
, and the initial relative frequency varied  . We observed a critical frequency, ρc, above which u1 eventually dominates. ρc was plotted for a range of toxicity (γA) and induction ability (l
A) values. As expected, ρc increased with both γA and l
A. Parameters: γB = 2, l
A = 40.
. We observed a critical frequency, ρc, above which u1 eventually dominates. ρc was plotted for a range of toxicity (γA) and induction ability (l
A) values. As expected, ρc increased with both γA and l
A. Parameters: γB = 2, l
A = 40.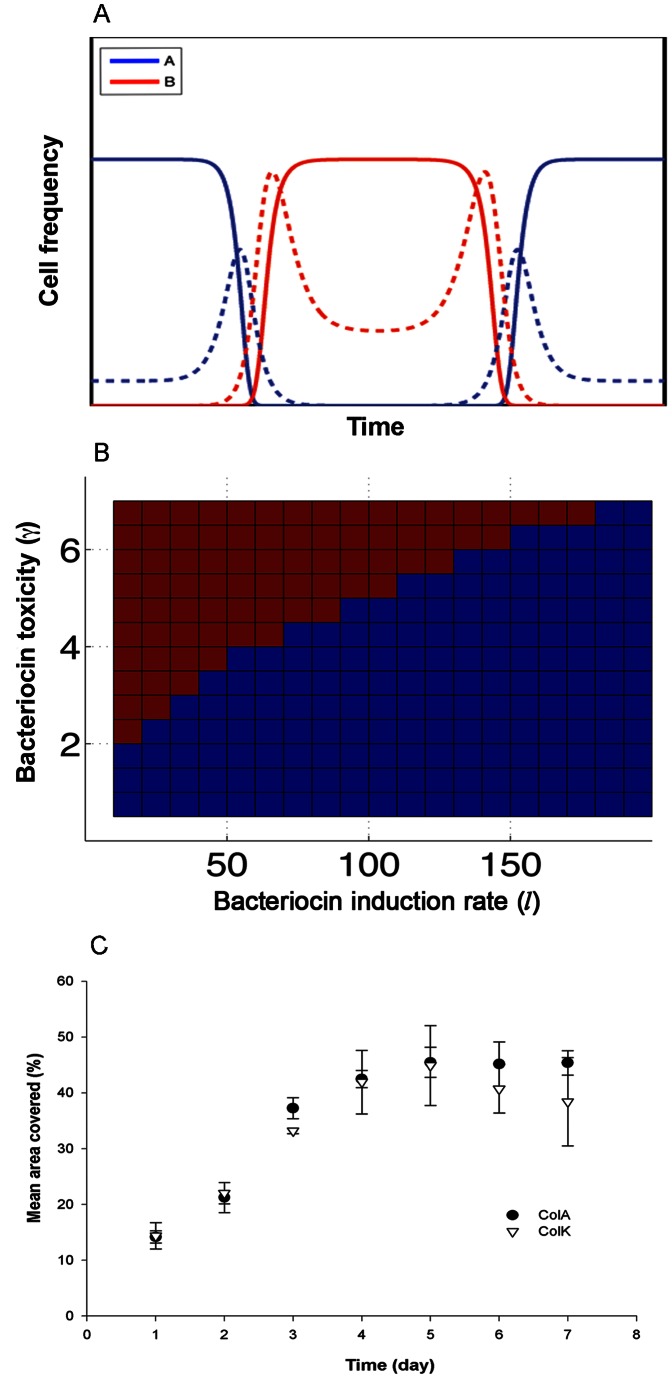
References
-
- de Boer W, Folman LB, Summerbell RC, Boddy L (2005) Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol Rev 29: 795–811. - PubMed
-
- Riley MA, Chavan MA (2007) Bacteriocins: Ecology and Evolution. Bacteriocins: Ecology and Evolution.
-
- Kerr B, Riley MA, Feldman MW, Bohannan BJ (2002) Local dispersal promotes biodiversity in a real-life game of rock-paper-scissors. Nature 418: 171–174. - PubMed
-
- Gordon DM, Riley MA (1999) A theoretical and empirical investigation of the invasion dynamics of colicinogeny. Microbiology 145: 655–661. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
