

Genetic dissection of ventral folding morphogenesis in mouse: embryonic visceral endoderm-supplied BMP2 positions head and heart
- PMID: 23706163
- PMCID: PMC3759531
- DOI: 10.1016/j.gde.2013.04.001
Genetic dissection of ventral folding morphogenesis in mouse: embryonic visceral endoderm-supplied BMP2 positions head and heart
Abstract
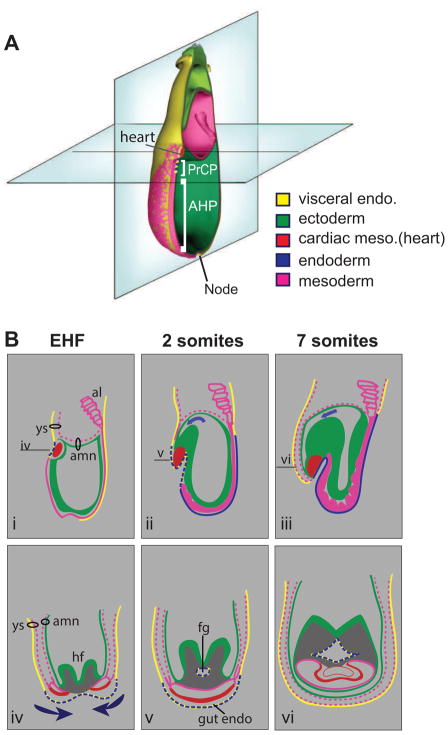
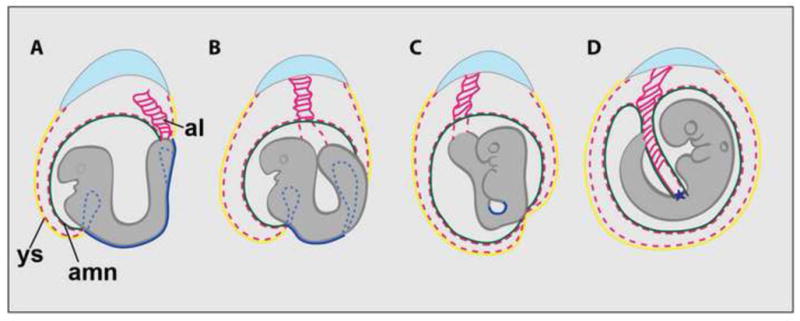
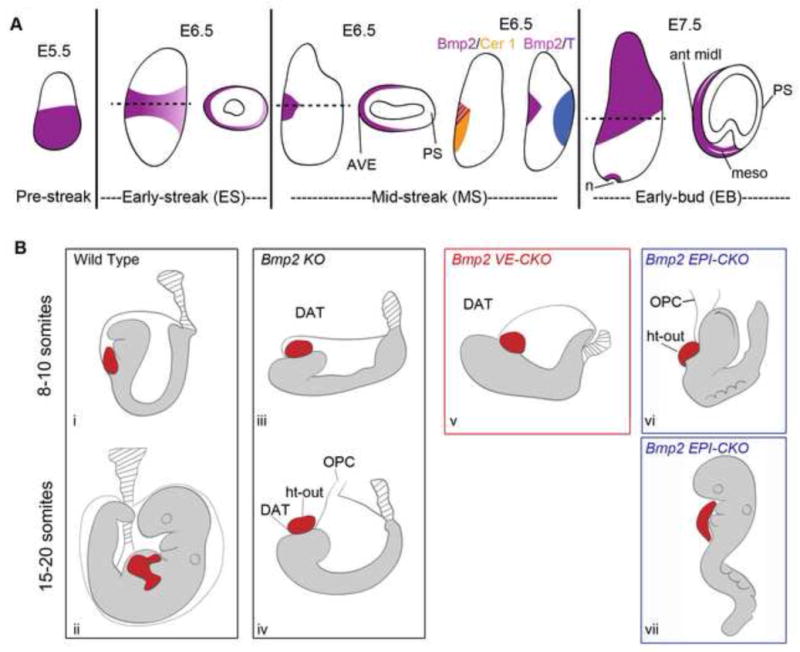
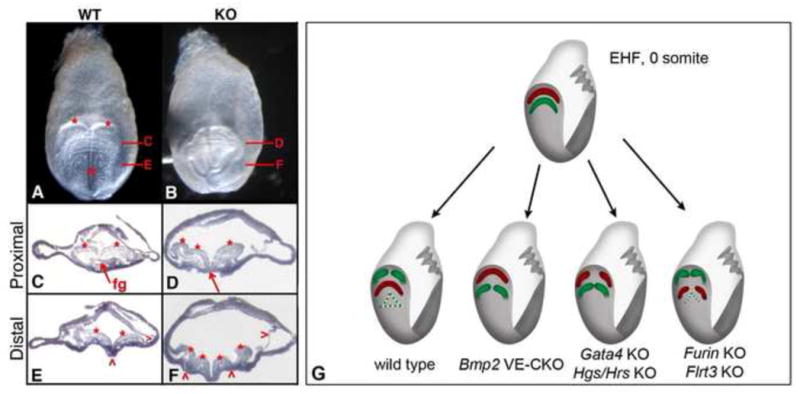
Ventral folding morphogenesis (VFM), a vital morphogenetic process in amniotes, mediates gut endoderm internalization, linear heart tube formation, ventral body wall closure and encasement of the fetus in extraembryonic membranes. Aberrant VFM underlies a number of birth defects such as gastroschisis and ectopia cordis in human and misplacement of head and heart in mouse. Recent cell lineage-specific mouse mutant analyses identified the Bone Morphogenetic Protein (BMP) pathway and Anterior Visceral Endoderm (AVE) as key regulators of anterior VFM. Loss of BMP2 expression solely from embryonic visceral endoderm (EmVE) and the AVE blocks formation of foregut invagination, and simultaneously, aberrantly positions the heart anterior/dorsal to the head, suggesting a mechanistic link between foregut and head/heart morphogenesis.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Sadler TW, Feldkamp ML. The embryology of body wall closure: relevance to gastroschisis and other ventral body wall defects. Am J Med Genet C Semin Med Genet. 2008;148C:180–185. - PubMed
-
- Williams T. Animal models of ventral body wall closure defects: a personal perspective on gastroschisis. Am J Med Genet C Semin Med Genet. 2008;148C:186–191. - PubMed
-
- Frolov P, Alali J, Klein MD. Clinical risk factors for gastroschisis and omphalocele in humans: a review of the literature. Pediatr Surg Int. 2010;26:1135–1148. - PubMed
-
- Kaufman MB. The Atlas of Mouse Development. London: Academic Press; 1992.
-
- Sadler TW. The embryologic origin of ventral body wall defects. Semin Pediatr Surg. 2010;19:209–214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
