

Architectural protein subclasses shape 3D organization of genomes during lineage commitment
- PMID: 23706625
- PMCID: PMC3712340
- DOI: 10.1016/j.cell.2013.04.053
Architectural protein subclasses shape 3D organization of genomes during lineage commitment
Abstract
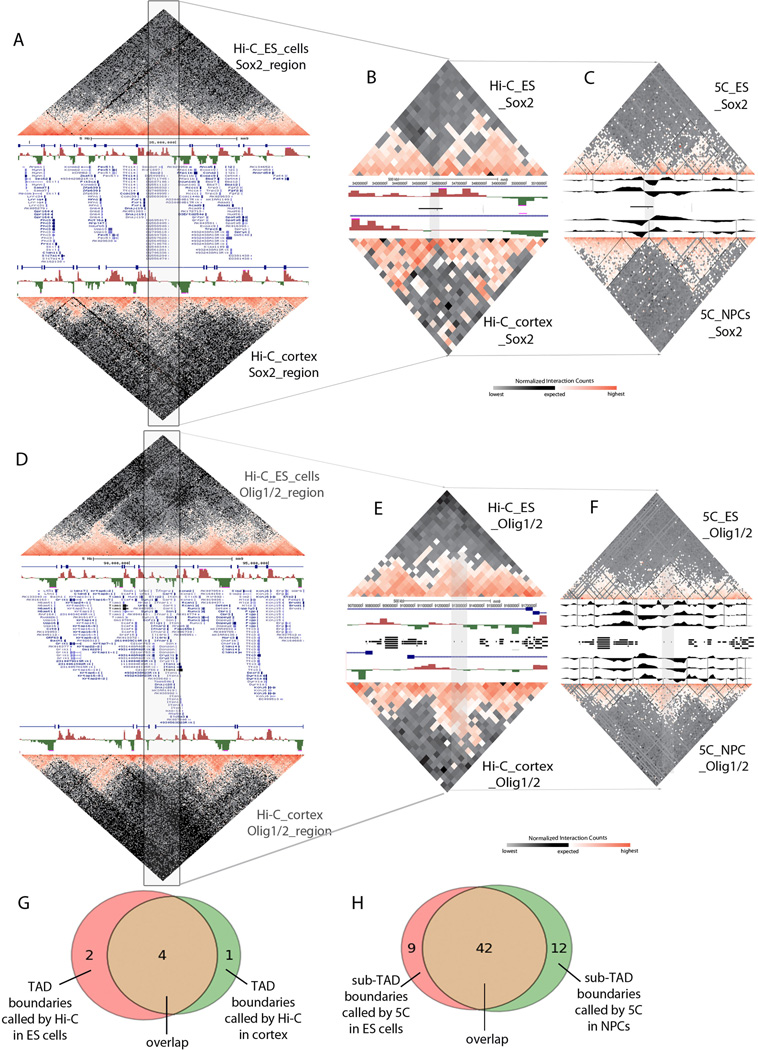
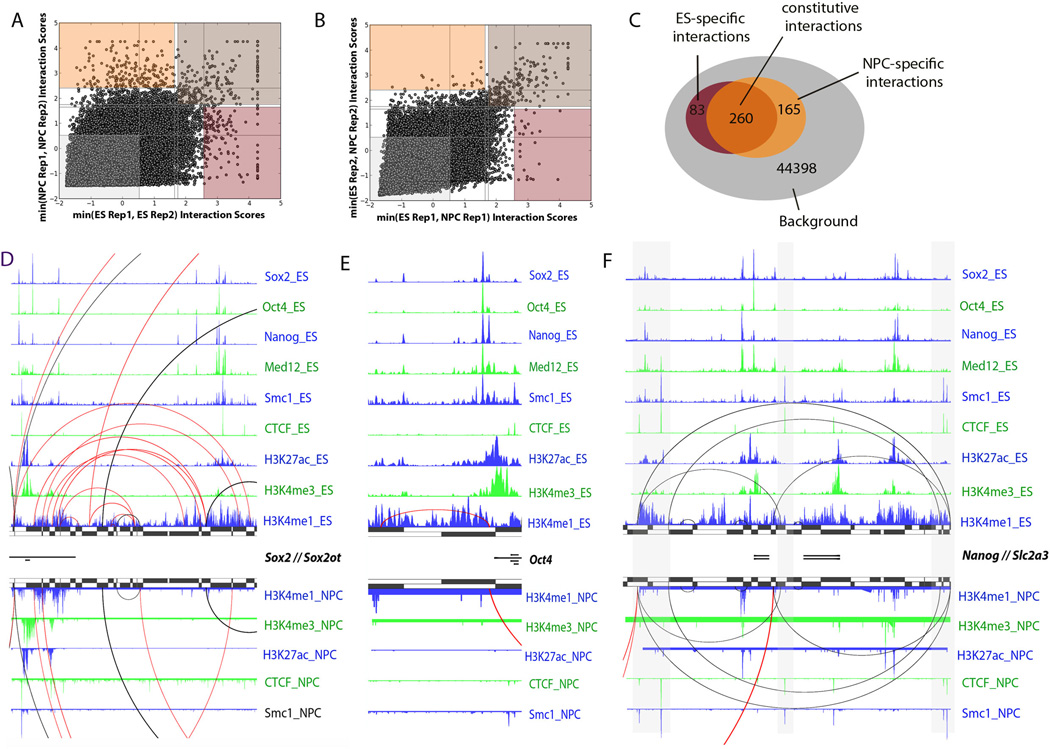
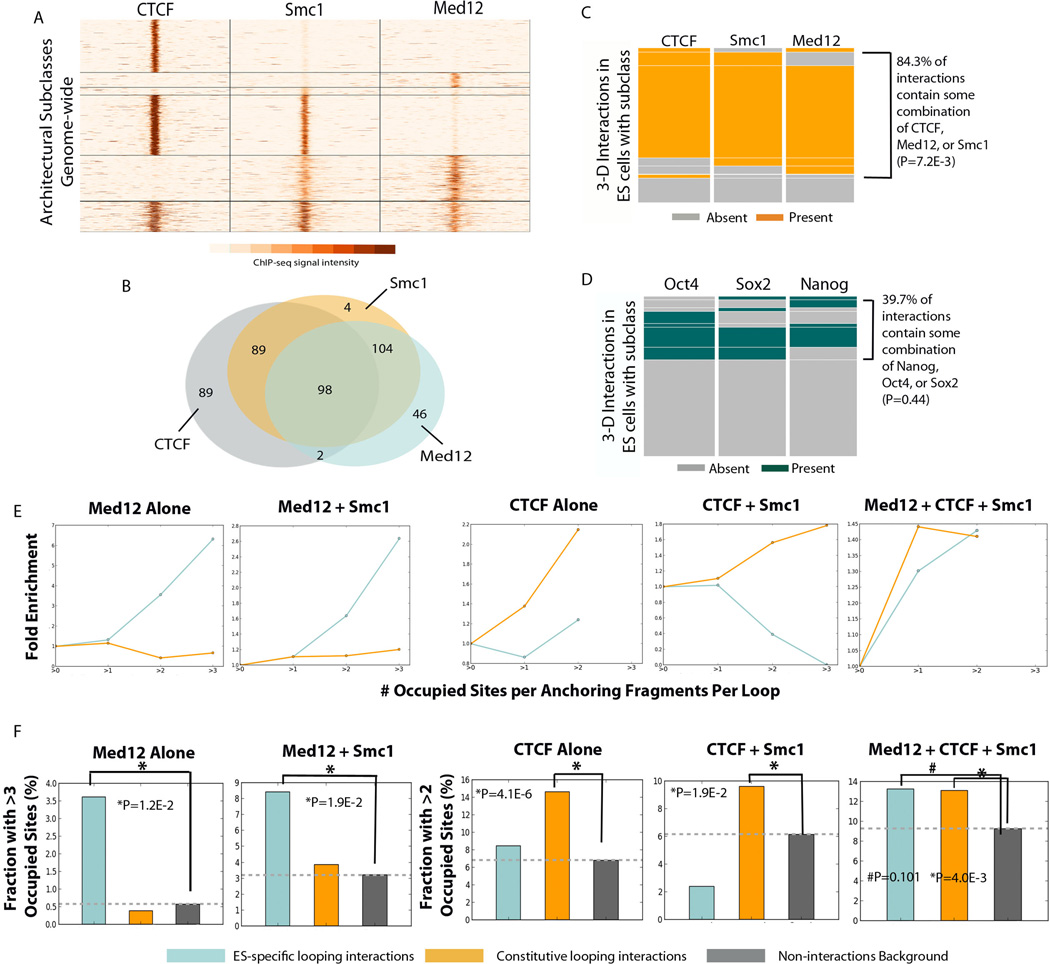
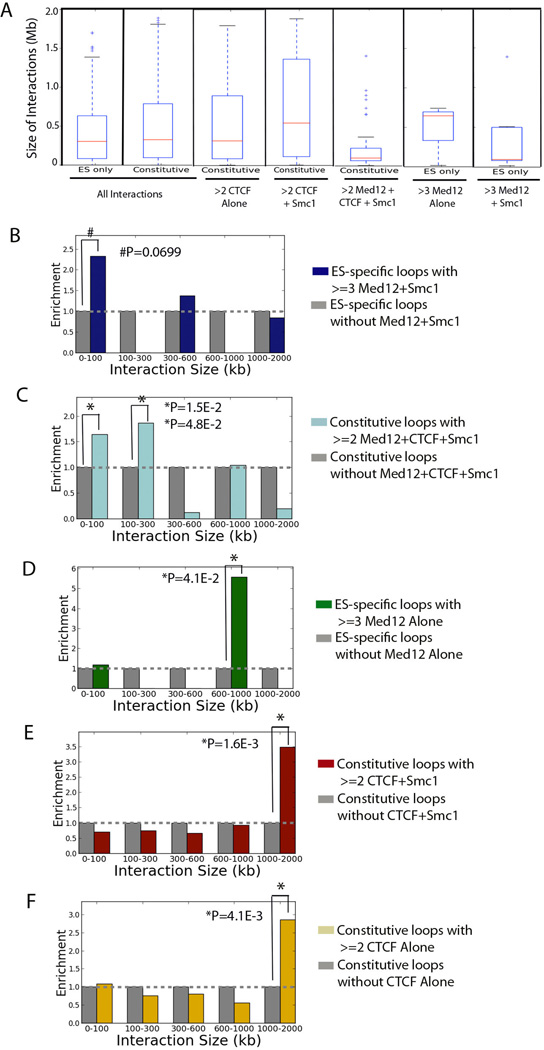
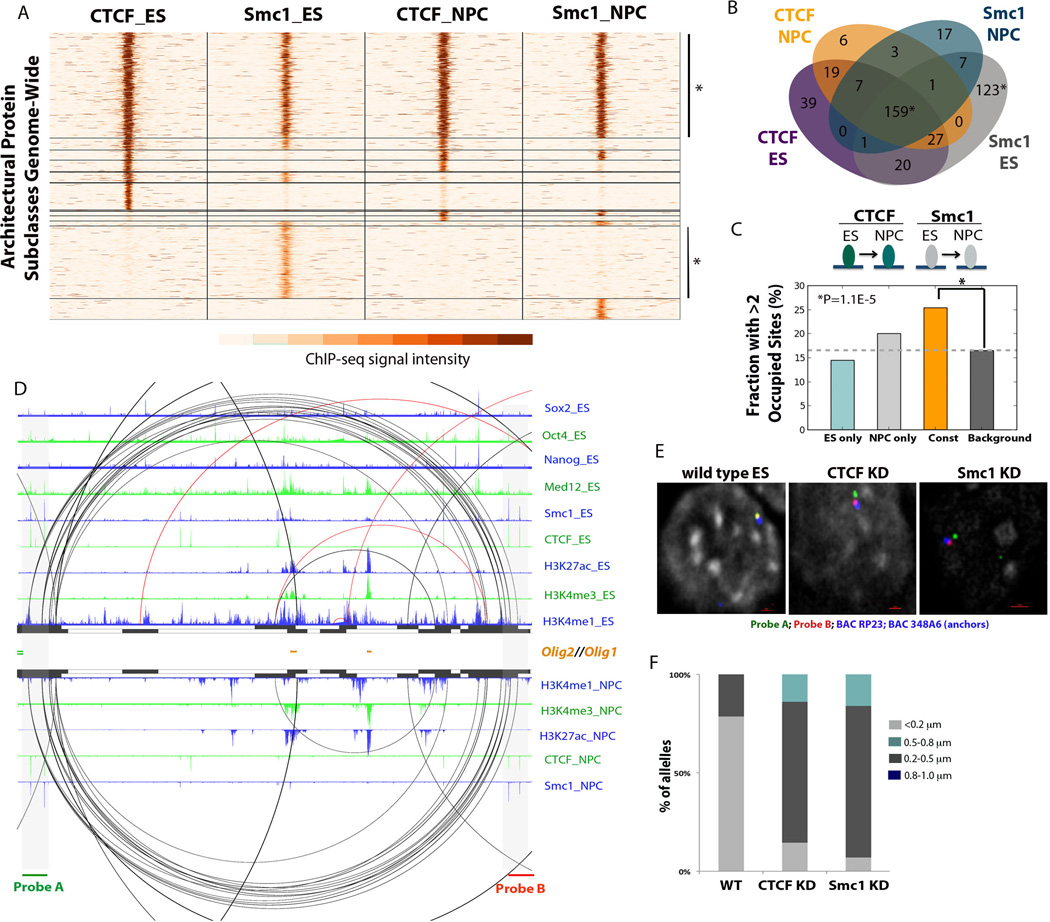
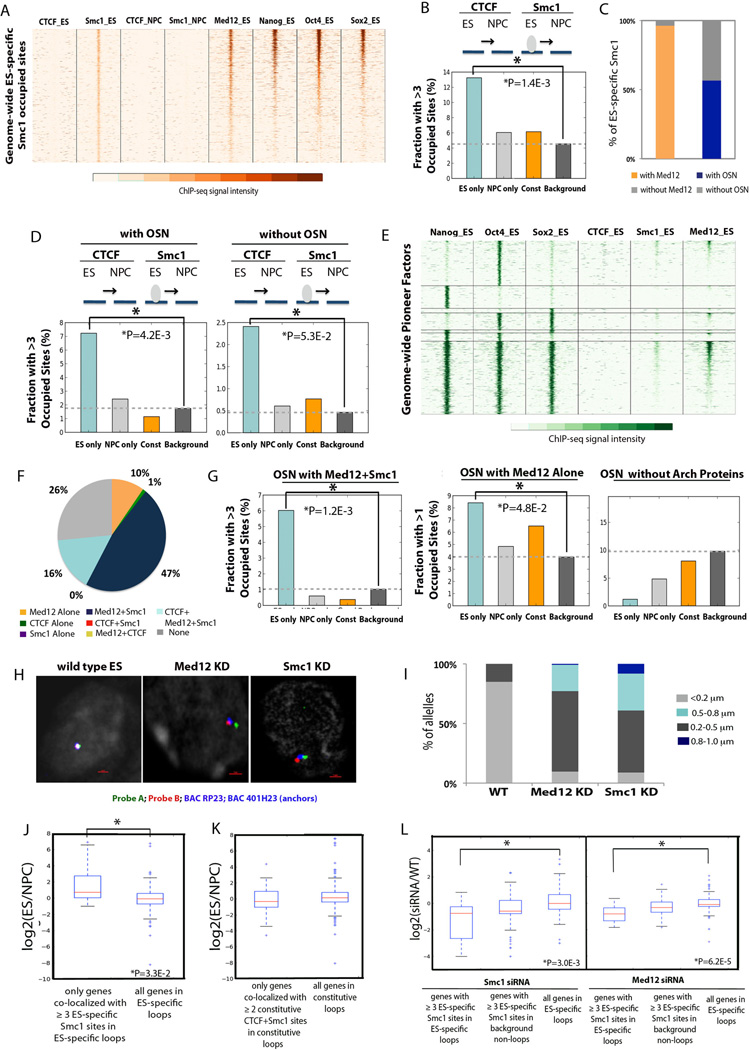
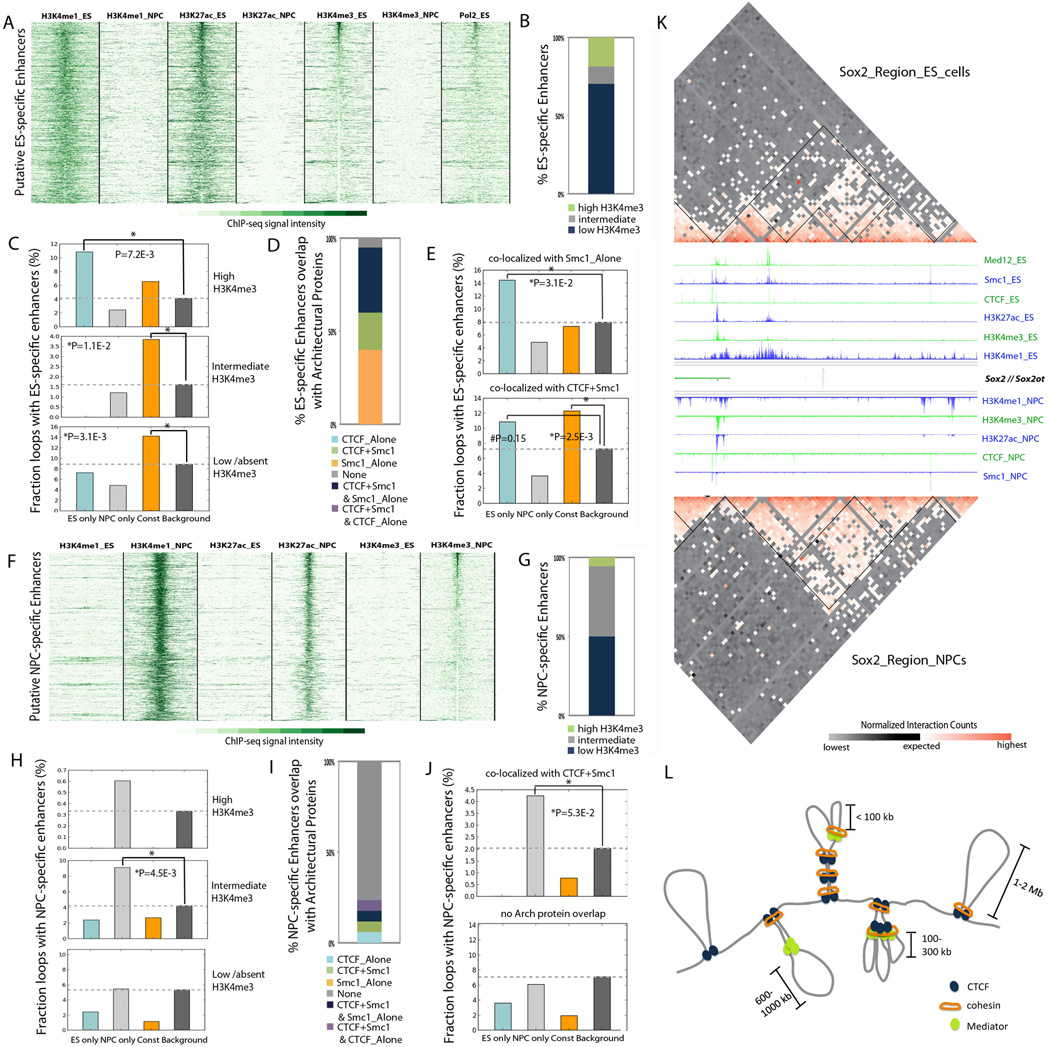
Understanding the topological configurations of chromatin may reveal valuable insights into how the genome and epigenome act in concert to control cell fate during development. Here, we generate high-resolution architecture maps across seven genomic loci in embryonic stem cells and neural progenitor cells. We observe a hierarchy of 3D interactions that undergo marked reorganization at the submegabase scale during differentiation. Distinct combinations of CCCTC-binding factor (CTCF), Mediator, and cohesin show widespread enrichment in chromatin interactions at different length scales. CTCF/cohesin anchor long-range constitutive interactions that might form the topological basis for invariant subdomains. Conversely, Mediator/cohesin bridge short-range enhancer-promoter interactions within and between larger subdomains. Knockdown of Smc1 or Med12 in embryonic stem cells results in disruption of spatial architecture and downregulation of genes found in cohesin-mediated interactions. We conclude that cell-type-specific chromatin organization occurs at the submegabase scale and that architectural proteins shape the genome in hierarchical length scales.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Chromatin meets its organizers.Cell. 2013 Jun 6;153(6):1187-9. doi: 10.1016/j.cell.2013.05.030. Cell. 2013. PMID: 23746835
-
The 3D genome shapes up for pluripotency.Cell Stem Cell. 2013 Jul 3;13(1):3-4. doi: 10.1016/j.stem.2013.06.013. Cell Stem Cell. 2013. PMID: 23827701
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 GM035463/GM/NIGMS NIH HHS/United States
- P01GM85354/GM/NIGMS NIH HHS/United States
- R01HG003143/HG/NHGRI NIH HHS/United States
- R01 HG003143/HG/NHGRI NIH HHS/United States
- P01 GM085354/GM/NIGMS NIH HHS/United States
- RC2HG005542/HG/NHGRI NIH HHS/United States
- RC2 HG005542/HG/NHGRI NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- R01 DK065806/DK/NIDDK NIH HHS/United States
- R01DK065806/DK/NIDDK NIH HHS/United States
- 5F32NS065603/NS/NINDS NIH HHS/United States
- F32 NS065603/NS/NINDS NIH HHS/United States
- R01GM035463/GM/NIGMS NIH HHS/United States
- T32 GM008490/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
