

Rhythmic modulation of the hematopoietic niche through neutrophil clearance
- PMID: 23706740
- PMCID: PMC4128329
- DOI: 10.1016/j.cell.2013.04.040
Rhythmic modulation of the hematopoietic niche through neutrophil clearance
Abstract
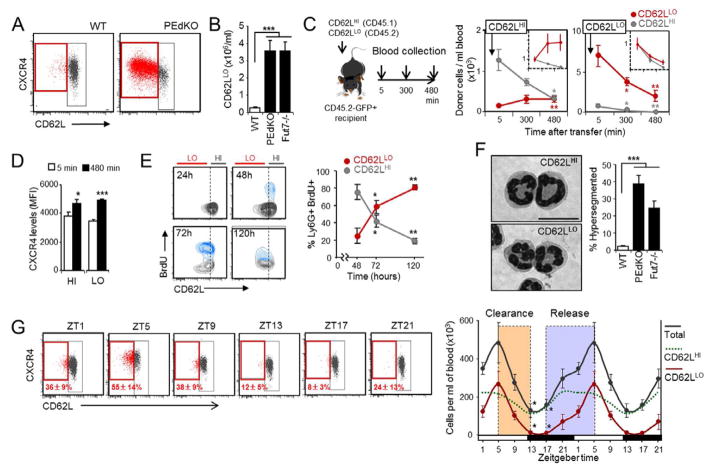
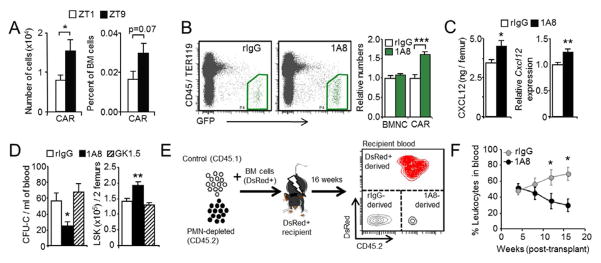
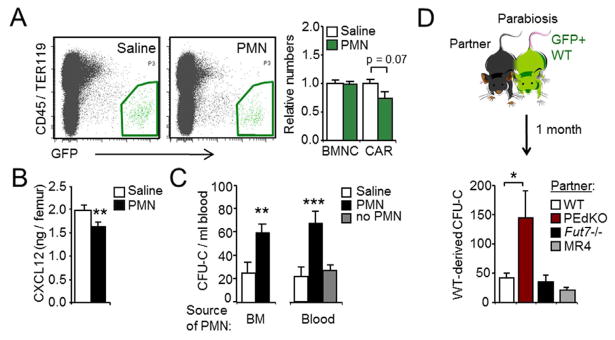
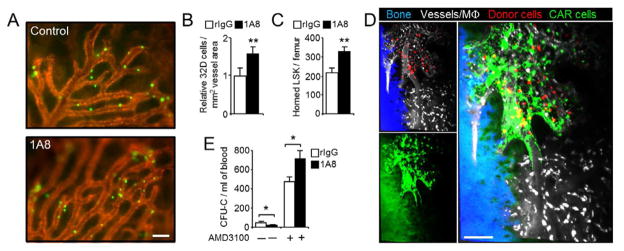
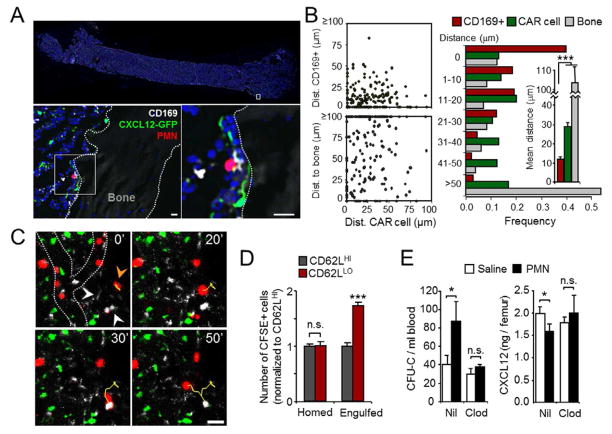
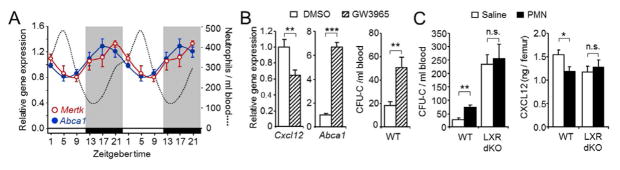
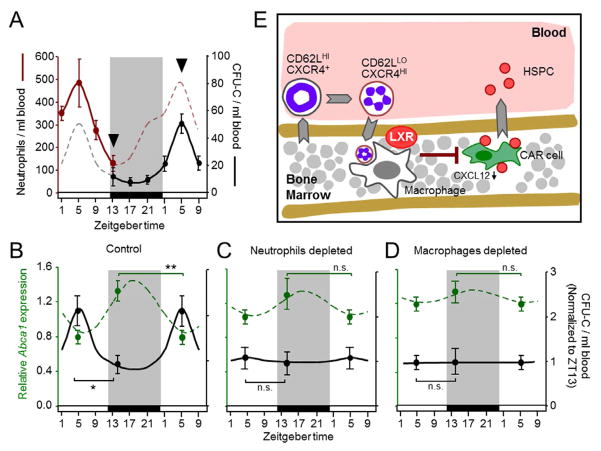
Unique among leukocytes, neutrophils follow daily cycles of release from and migration back into the bone marrow, where they are eliminated. Because removal of dying cells generates homeostatic signals, we explored whether neutrophil elimination triggers circadian events in the steady state. Here, we report that the homeostatic clearance of neutrophils provides cues that modulate the physiology of the bone marrow. We identify a population of CD62L(LO) CXCR4(HI) neutrophils that have "aged" in the circulation and are eliminated at the end of the resting period in mice. Aged neutrophils infiltrate the bone marrow and promote reductions in the size and function of the hematopoietic niche. Modulation of the niche depends on macrophages and activation of cholesterol-sensing nuclear receptors and is essential for the rhythmic egress of hematopoietic progenitors into the circulation. Our results unveil a process that synchronizes immune and hematopoietic rhythms and expand the ascribed functions of neutrophils beyond inflammation. PAPERFLICK:
Copyright © 2013 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Comment in
-
Haematopoiesis: neutrophil clearance keeps the beat.Nat Rev Immunol. 2013 Jul;13(7):470. doi: 10.1038/nri3483. Epub 2013 Jun 14. Nat Rev Immunol. 2013. PMID: 23765058 No abstract available.
References
-
- Adams GB, Martin RP, Alley IR, Chabner KT, Cohen KS, Calvi LM, Kronenberg HM, Scadden DT. Therapeutic targeting of a stem cell niche. Nat Biotechnol. 2007;25:238–243. - PubMed
-
- Basu S, Hodgson G, Katz M, Dunn AR. Evaluation of role of G-CSF in the production, survival, and release of neutrophils from bone marrow into circulation. Blood. 2002;100:854–861. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
