

RPA coordinates DNA end resection and prevents formation of DNA hairpins
- PMID: 23706822
- PMCID: PMC3855855
- DOI: 10.1016/j.molcel.2013.04.032
RPA coordinates DNA end resection and prevents formation of DNA hairpins
Abstract
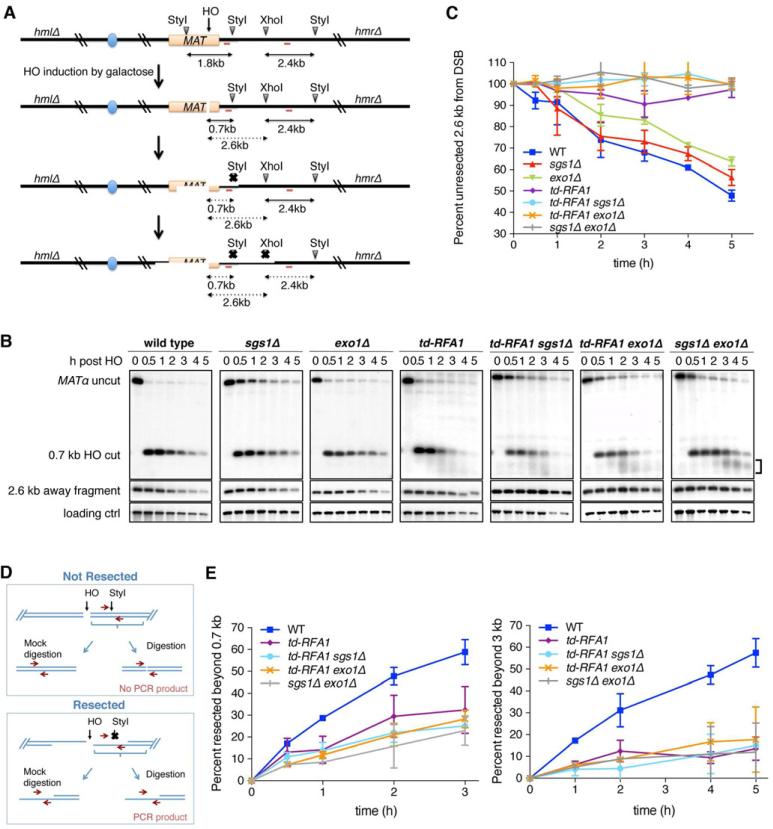
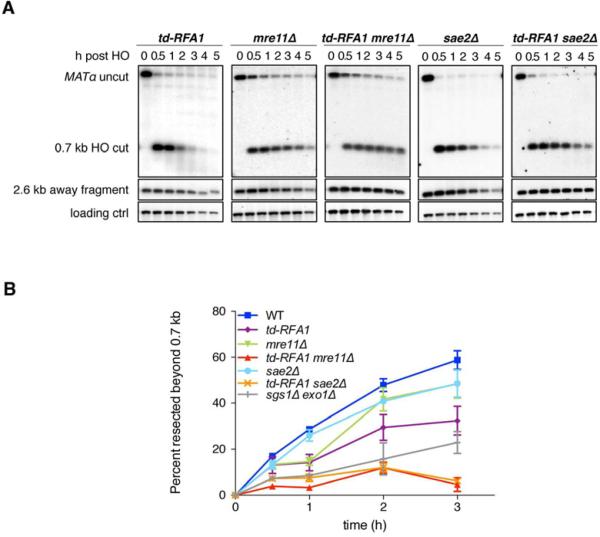
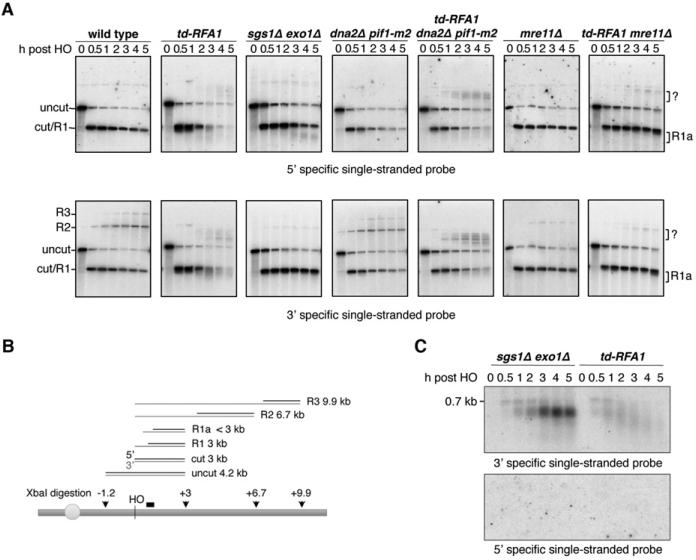
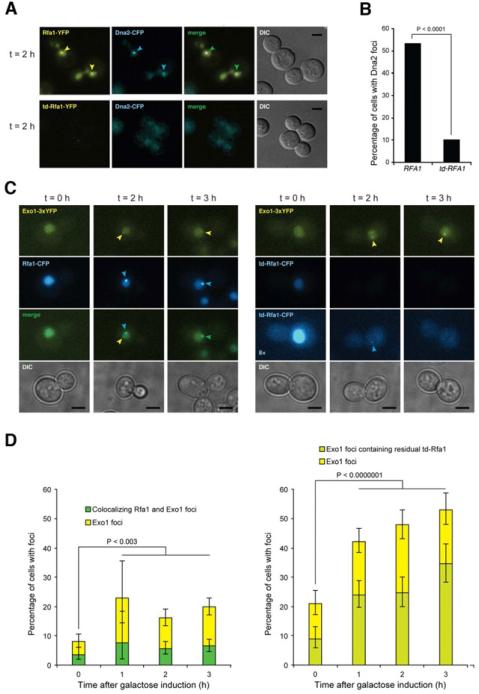
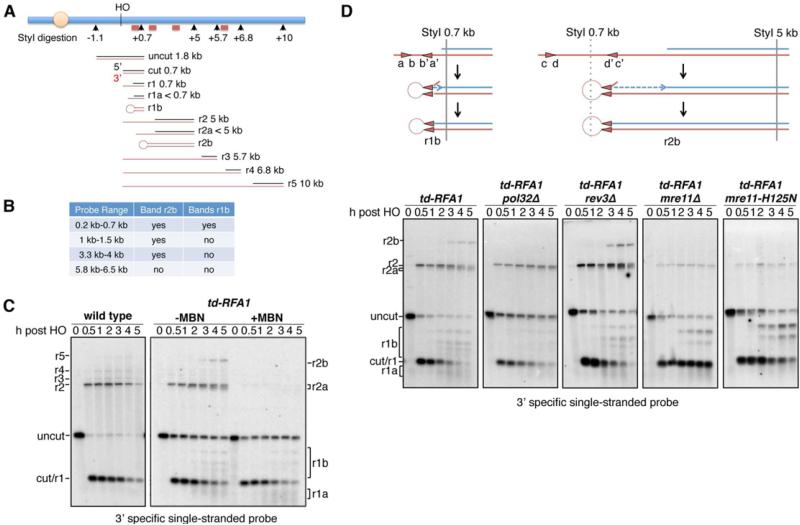
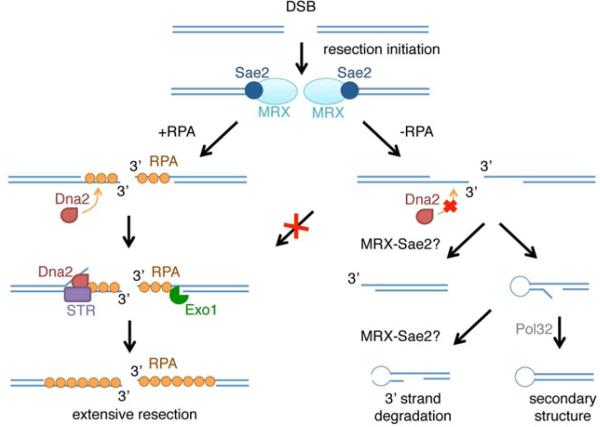
Replication protein A (RPA) is an essential eukaryotic single-stranded DNA binding protein with a central role in DNA metabolism. RPA directly participates in DNA double-strand break repair by stimulating 5'-3' end resection by the Sgs1/BLM helicase and Dna2 endonuclease in vitro. Here we investigated the role of RPA in end resection in vivo, using a heat-inducible degron system that allows rapid conditional depletion of RPA in Saccharomyces cerevisiae. We found that RPA depletion eliminated both the Sgs1-Dna2- and Exo1-dependent extensive resection pathways and synergized with mre11Δ to prevent end resection. The short single-stranded DNA tails formed in the absence of RPA were unstable due to 3' strand loss and the formation of fold-back hairpin structures that required resection initiation and Pol32-dependent DNA synthesis. Thus, RPA is required to generate ssDNA, and also to protect ssDNA from degradation and inappropriate annealing that could lead to genome rearrangements.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Amberg DC, Burke DJ, Strathern JN. Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual. Cold spring Harbor Laboratory Press; 2005.
-
- Bae SH, Seo YS. Characterization of the enzymatic properties of the yeast dna2 Helicase/endonuclease suggests a new model for Okazaki fragment processing. J Biol. Chem. 2000;275:38022–38031. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
