

The NuRD complex cooperates with DNMTs to maintain silencing of key colorectal tumor suppressor genes
- PMID: 23708667
- PMCID: PMC3883927
- DOI: 10.1038/onc.2013.178
The NuRD complex cooperates with DNMTs to maintain silencing of key colorectal tumor suppressor genes
Abstract
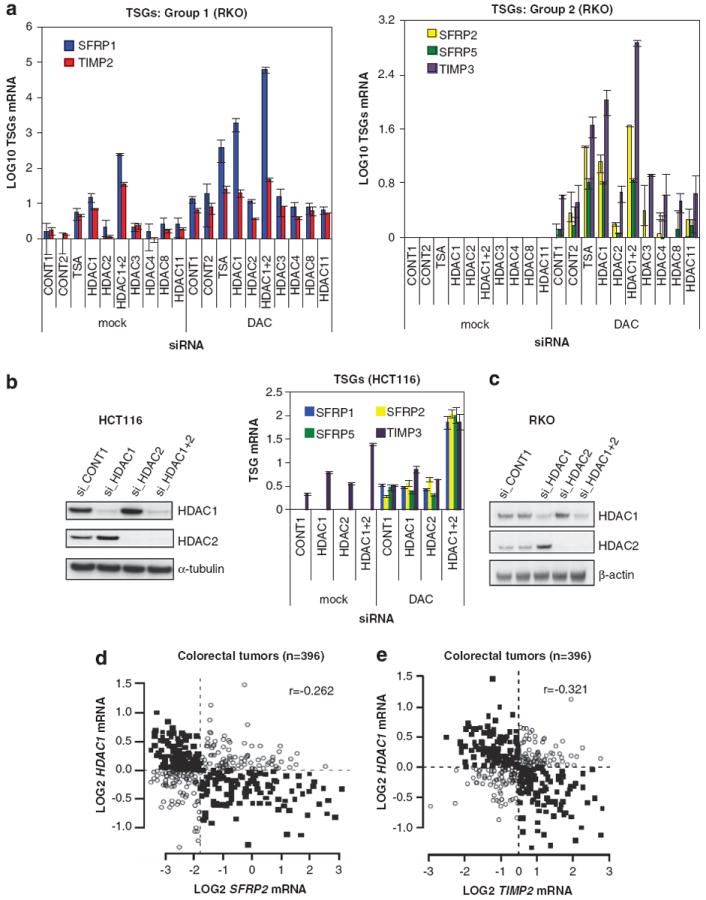
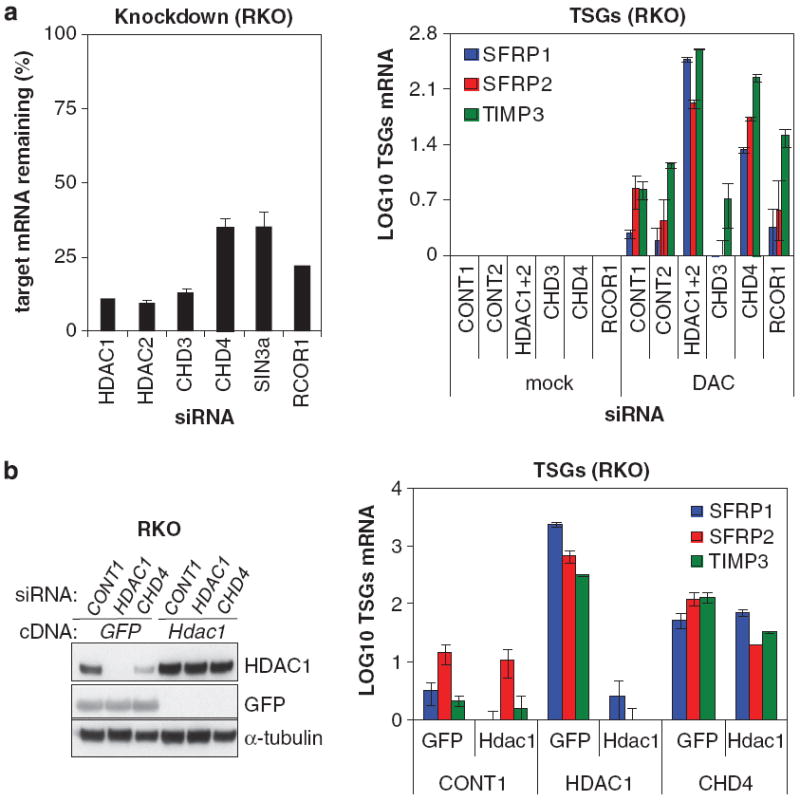
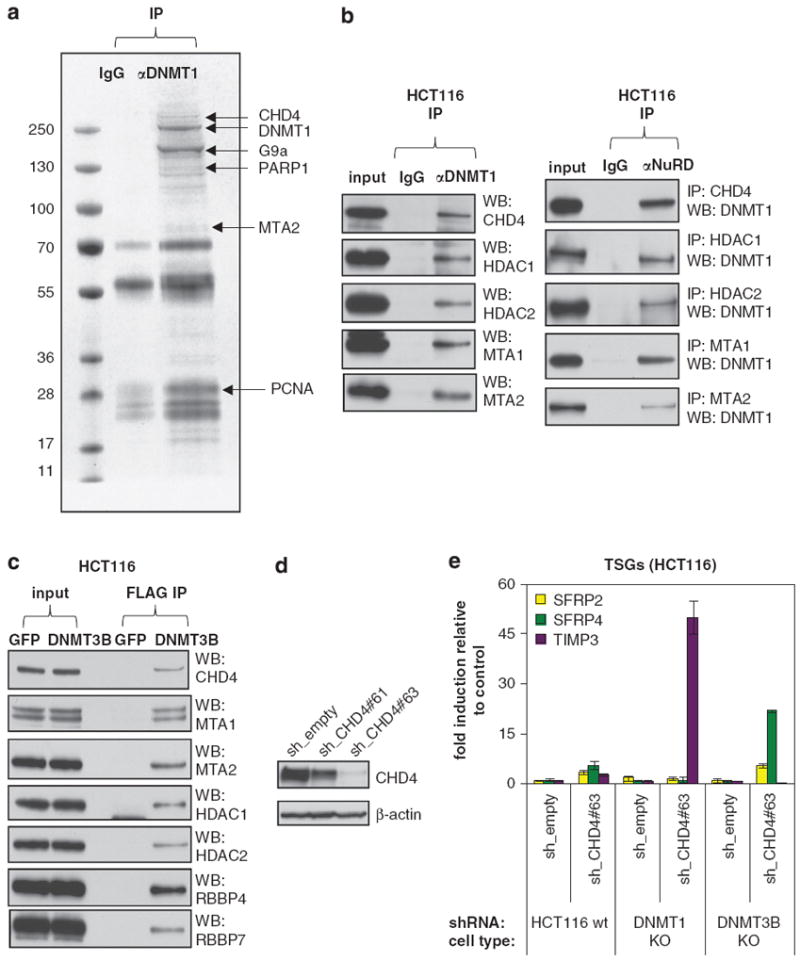
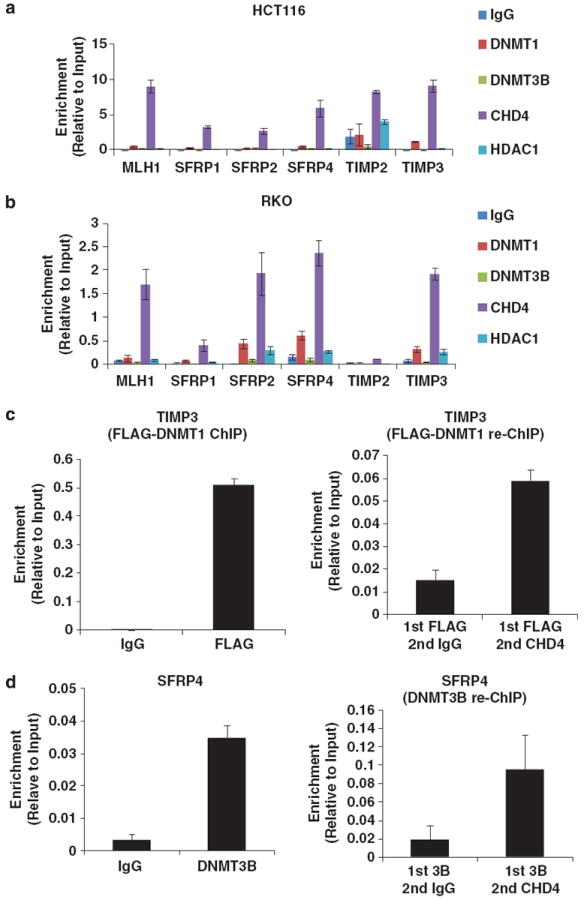
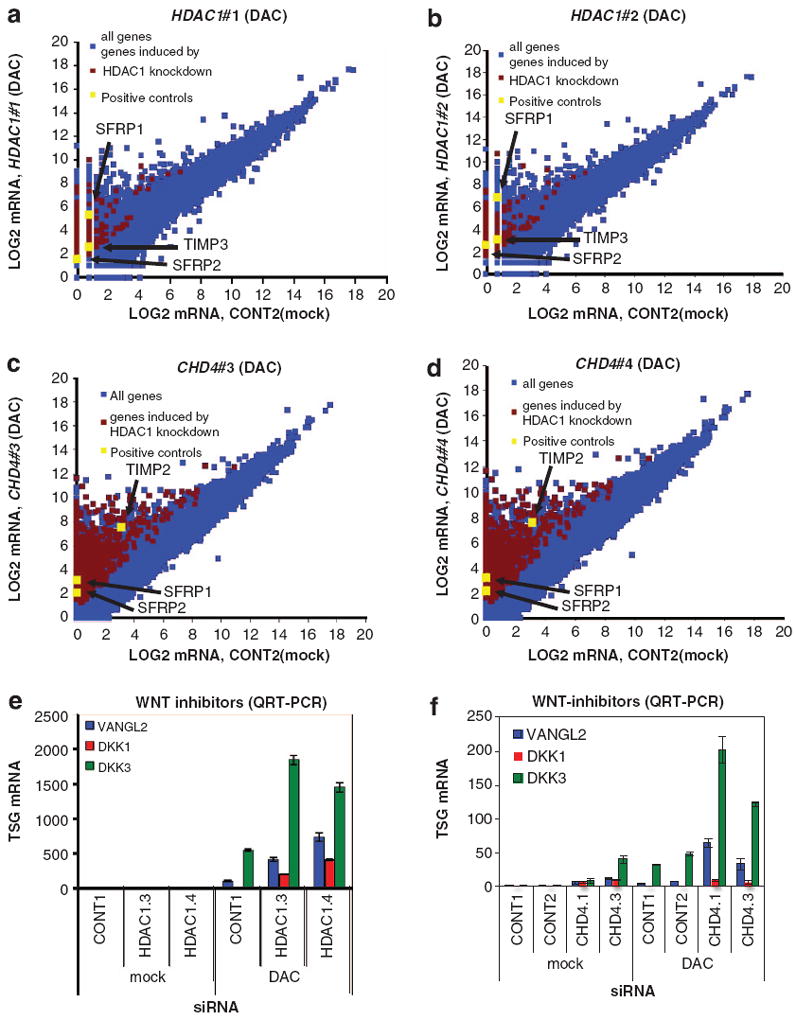
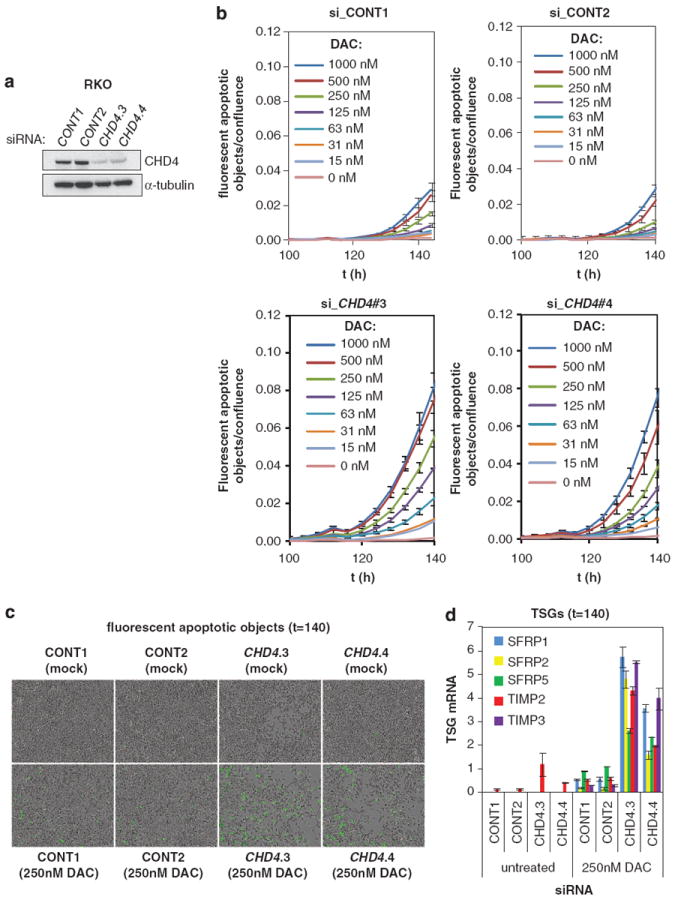
Many tumor suppressor genes (TSGs) are silenced through synergistic layers of epigenetic regulation including abnormal DNA hypermethylation of promoter CpG islands, repressive chromatin modifications and enhanced nucleosome deposition over transcription start sites. The protein complexes responsible for silencing of many of such TSGs remain to be identified. Our previous work demonstrated that multiple silenced TSGs in colorectal cancer cells can be partially reactivated by DNA demethylation in cells disrupted for the DNA methyltransferases 1 and 3B (DNMT1 and 3B) or by DNMT inhibitors (DNMTi). Herein, we used proteomic and functional genetic approaches to identify additional proteins that cooperate with DNMTs in silencing these key silenced TSGs in colon cancer cells. We discovered that DNMTs and the core components of the NuRD (Mi-2/nucleosome remodeling and deacetylase) nucleosome remodeling complex, chromo domain helicase DNA-binding protein 4 (CHD4) and histone deacetylase 1 (HDAC1) occupy the promoters of several of these hypermethylated TSGs and physically and functionally interact to maintain their silencing. Consistent with this, we find an inverse relationship between expression of HDAC1 and 2 and these TSGs in a large panel of primary colorectal tumors. We demonstrate that DNMTs and NuRD cooperate to maintain the silencing of several negative regulators of the WNT and other signaling pathways. We find that depletion of CHD4 is synergistic with DNMT inhibition in reducing the viability of colon cancer cells in correlation with reactivation of TSGs, suggesting that their combined inhibition may be beneficial for the treatment of colon cancer. Since CHD4 has ATPase activity, our data identify CHD4 as a potentially novel drug target in cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
