

Two salt bridges differentially contribute to the maintenance of cystic fibrosis transmembrane conductance regulator (CFTR) channel function
- PMID: 23709221
- PMCID: PMC3711338
- DOI: 10.1074/jbc.M113.476226
Two salt bridges differentially contribute to the maintenance of cystic fibrosis transmembrane conductance regulator (CFTR) channel function
Abstract
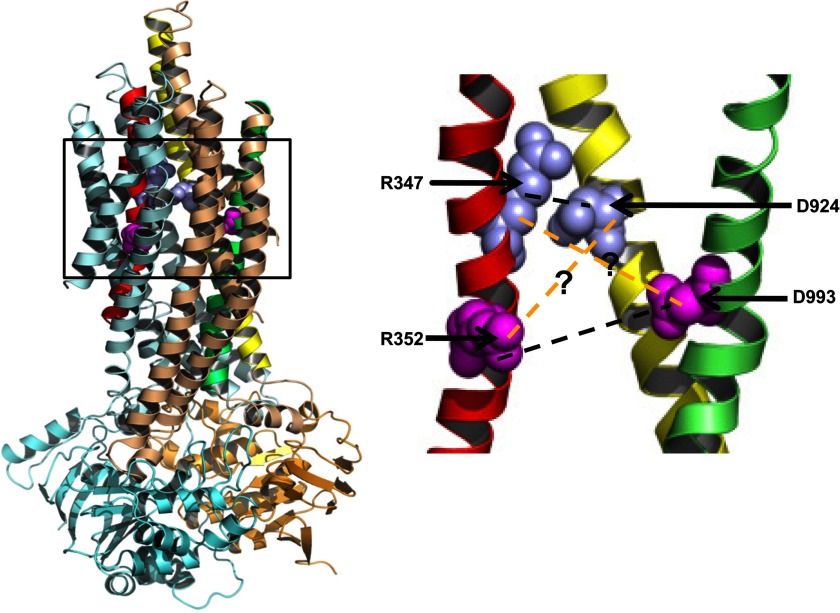
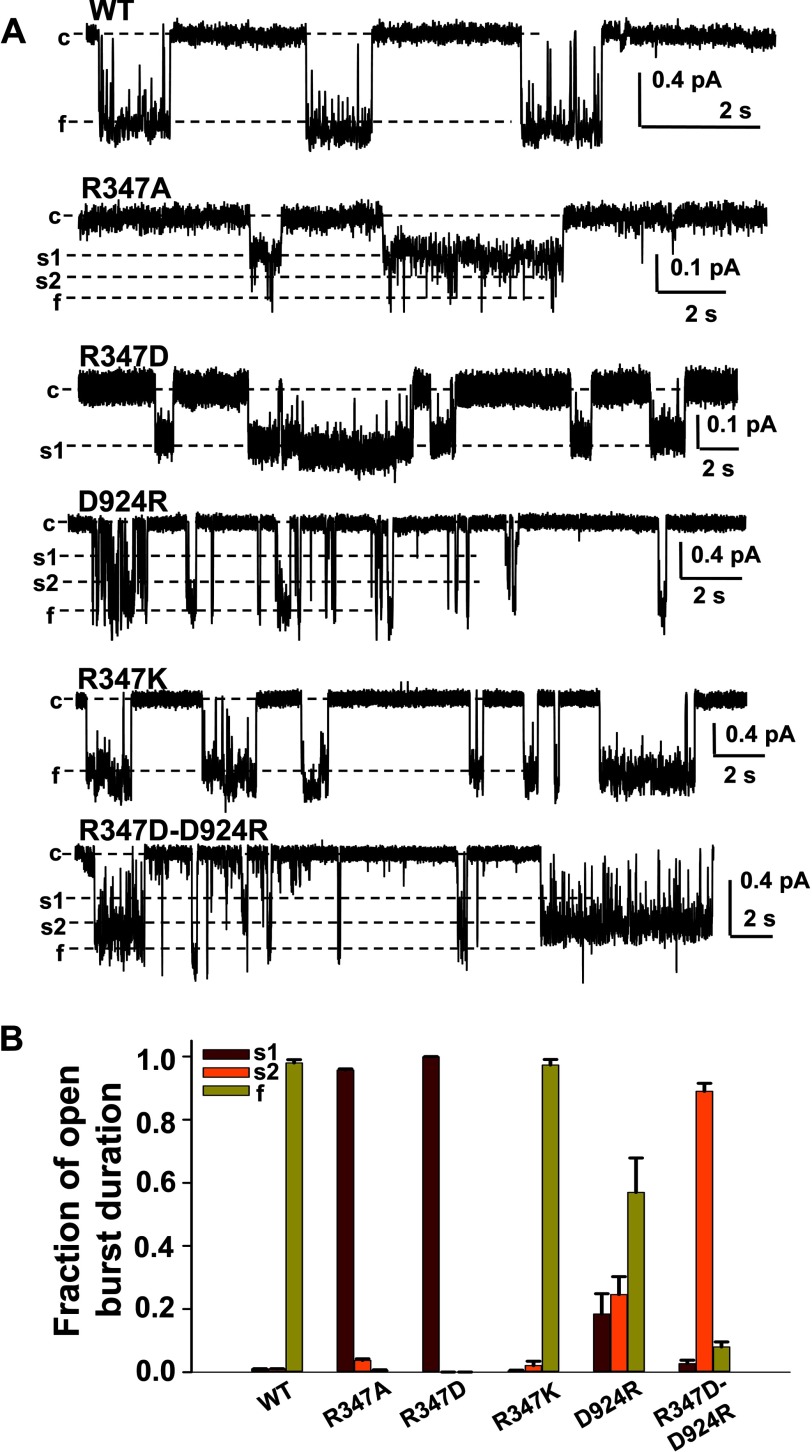
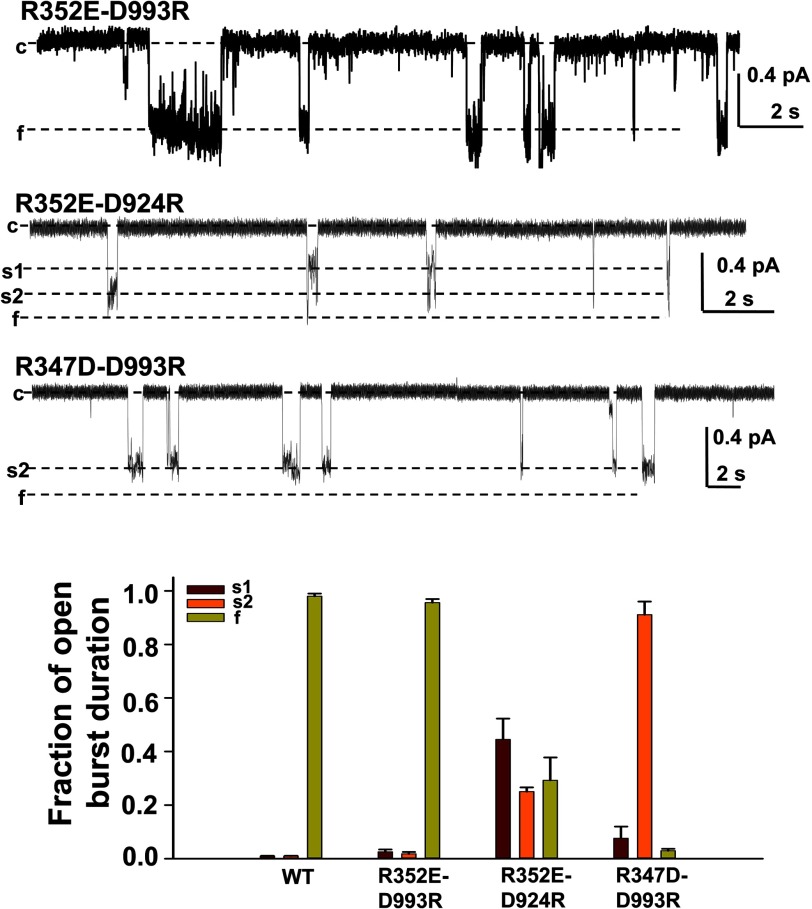
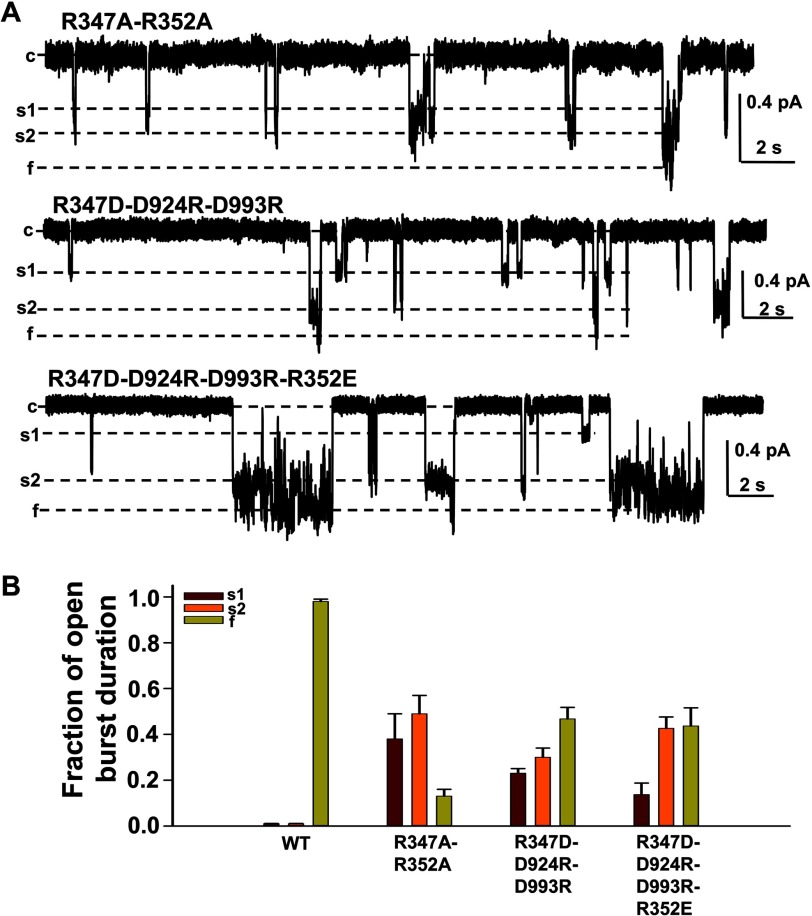
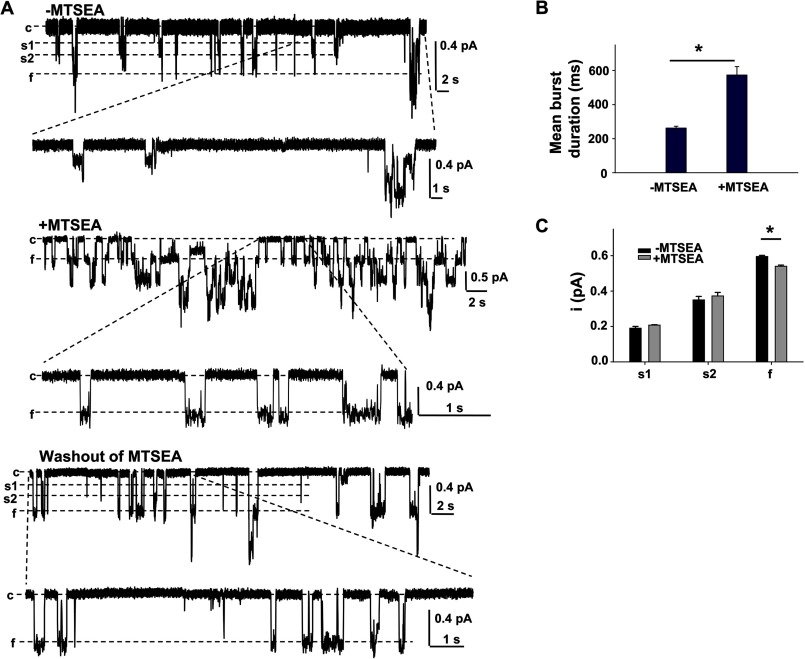
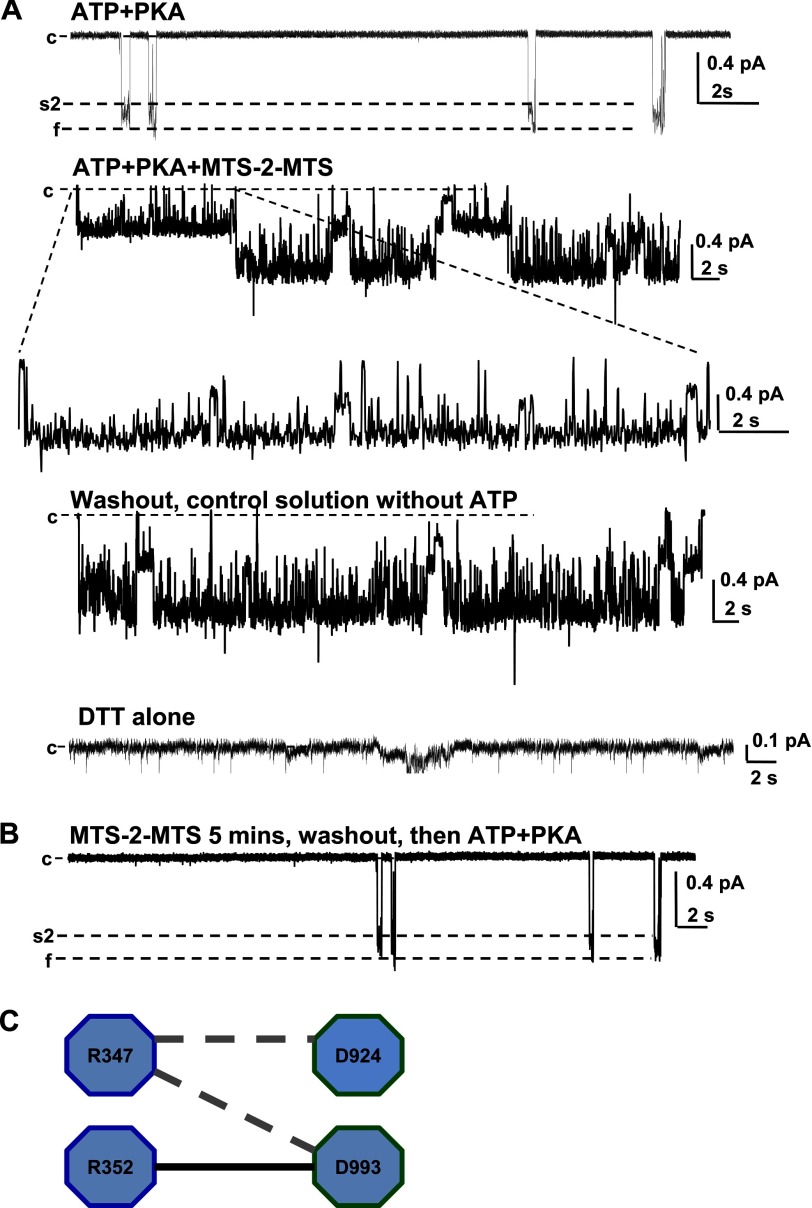
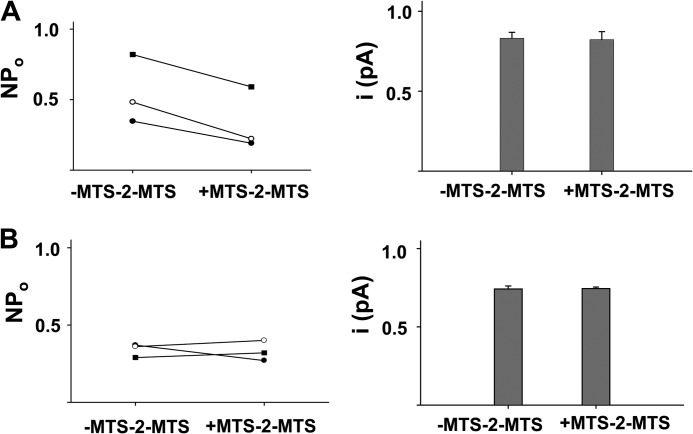
Previous studies have identified two salt bridges in human CFTR chloride ion channels, Arg(352)-Asp(993) and Arg(347)-Asp(924), that are required for normal channel function. In the present study, we determined how the two salt bridges cooperate to maintain the open pore architecture of CFTR. Our data suggest that Arg(347) not only interacts with Asp(924) but also interacts with Asp(993). The tripartite interaction Arg(347)-Asp(924)-Asp(993) mainly contributes to maintaining a stable s2 open subconductance state. The Arg(352)-Asp(993) salt bridge, in contrast, is involved in stabilizing both the s2 and full (f) open conductance states, with the main contribution being to the f state. The s1 subconductance state does not require either salt bridge. In confirmation of the role of Arg(352) and Asp(993), channels bearing cysteines at these sites could be latched into a full open state using the bifunctional cross-linker 1,2-ethanediyl bismethanethiosulfonate, but only when applied in the open state. Channels remained latched open even after washout of ATP. The results suggest that these interacting residues contribute differently to stabilizing the open pore in different phases of the gating cycle.
Keywords: ABC Transporter; CFTR; Gating; MTS Reagents; Oocyte; Pore; Salt Bridge; Xenopus.
Figures
References
-
- Alexander C., Ivetac A., Liu X., Norimatsu Y., Serrano J. R., Landstrom A., Sansom M., Dawson D. C. (2009) Cystic fibrosis transmembrane conductance regulator. Using differential reactivity toward channel-permeant and channel-impermeant thiol-reactive probes to test a molecular model for the pore. Biochemistry 48, 10078–10088 - PMC - PubMed
-
- Riordan J. R., Rommens J. M., Kerem B., Alon N., Rozmahel R., Grzelczak Z., Zielenski J., Lok S., Plavsic N., Chou J. L. (1989) Identification of the cystic fibrosis gene. Cloning and characterization of complementary DNA. Science 245, 1066–1073 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
