

Biochemical and structural characterization of the ubiquitin-conjugating enzyme UBE2W reveals the formation of a noncovalent homodimer
- PMID: 23709311
- PMCID: PMC3758794
- DOI: 10.1007/s12013-013-9633-5
Biochemical and structural characterization of the ubiquitin-conjugating enzyme UBE2W reveals the formation of a noncovalent homodimer
Abstract
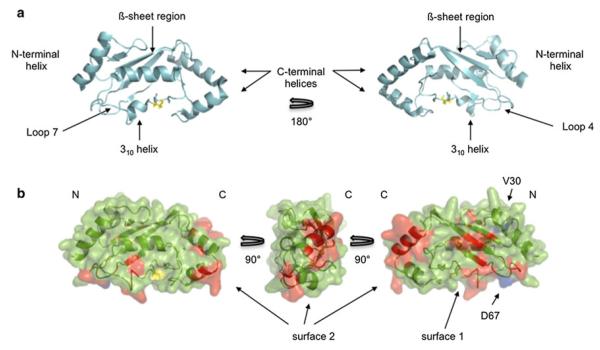
The biochemical and structural characterization of ubiquitin-conjugating enzymes (E2s) over the past 30 years has fostered important insights into ubiquitin transfer mechanisms. Although many of these enzymes share high sequence and structural conservation, their functional roles in the cell are decidedly diverse. Here, we report that the mono-ubiquitinating E2 UBE2W forms a homodimer using two distinct protein surfaces. Dimerization is primarily driven by residues in the ß-sheet region and Loops 4 and 7 of the catalytic domain. Mutation of two residues in the catalytic domain of UBE2W is capable of disrupting UBE2W homodimer formation, however, we find that dimerization of this E2 is not required for its ubiquitin transfer activity. In addition, residues in the C-terminal region, although not compulsory for the dimerization of UBE2W, play an ancillary role in the dimer interface. In all current E2 structures, the C-terminal helix of the UBC domain is at least 15Å away from the primary dimerization surface shown here for UBE2W. This leads to the proposal that the C-terminal region of UBE2W adopts a noncanonical position that places it closer to the UBC ß-sheet, providing the first indication that at least some E2s adopt C-terminal conformations different from the canonical structures observed to date.
Figures


Similar articles
-
Biochemical and structural characterization of a novel ubiquitin-conjugating enzyme E2 from Agrocybe aegeria reveals Ube2w family-specific properties.Sci Rep. 2015 Nov 3;5:16056. doi: 10.1038/srep16056. Sci Rep. 2015. PMID: 26525192 Free PMC article.
-
The ubiquitin-conjugating enzyme (E2) Ube2w ubiquitinates the N terminus of substrates.J Biol Chem. 2013 Jun 28;288(26):18784-8. doi: 10.1074/jbc.C113.477596. Epub 2013 May 21. J Biol Chem. 2013. PMID: 23696636 Free PMC article.
-
Solution structure of the flexible class II ubiquitin-conjugating enzyme Ubc1 provides insights for polyubiquitin chain assembly.J Biol Chem. 2004 Nov 5;279(45):47139-47. doi: 10.1074/jbc.M409576200. Epub 2004 Aug 24. J Biol Chem. 2004. PMID: 15328341
-
The family of ubiquitin-conjugating enzymes (E2s): deciding between life and death of proteins.FASEB J. 2010 Apr;24(4):981-93. doi: 10.1096/fj.09-136259. Epub 2009 Nov 25. FASEB J. 2010. PMID: 19940261 Review.
-
Ubiquitin-conjugating enzyme E2C: a potential cancer biomarker.Int J Biochem Cell Biol. 2014 Feb;47:113-7. doi: 10.1016/j.biocel.2013.11.023. Epub 2013 Dec 17. Int J Biochem Cell Biol. 2014. PMID: 24361302 Review.
Cited by
-
Ubiquitination is required for the initial removal of paternal organelles in C. elegans.Dev Biol. 2019 Sep 15;453(2):168-179. doi: 10.1016/j.ydbio.2019.05.015. Epub 2019 May 30. Dev Biol. 2019. PMID: 31153831 Free PMC article.
-
UBE2W Improves the Experimental Colitis by Inhibiting the NF-κB Signaling Pathway.Dig Dis Sci. 2022 Dec;67(12):5529-5539. doi: 10.1007/s10620-022-07453-4. Epub 2022 Mar 21. Dig Dis Sci. 2022. PMID: 35314916
-
Intrinsic disorder drives N-terminal ubiquitination by Ube2w.Nat Chem Biol. 2015 Jan;11(1):83-9. doi: 10.1038/nchembio.1700. Epub 2014 Dec 1. Nat Chem Biol. 2015. PMID: 25436519 Free PMC article.
-
Protein disorder: wagging a tail at ubiquitin.Nat Chem Biol. 2015 Jan;11(1):7-8. doi: 10.1038/nchembio.1716. Epub 2014 Dec 17. Nat Chem Biol. 2015. PMID: 25517385 No abstract available.
-
Loss of the Ubiquitin-conjugating Enzyme UBE2W Results in Susceptibility to Early Postnatal Lethality and Defects in Skin, Immune, and Male Reproductive Systems.J Biol Chem. 2016 Feb 5;291(6):3030-42. doi: 10.1074/jbc.M115.676601. Epub 2015 Nov 24. J Biol Chem. 2016. PMID: 26601958 Free PMC article.
References
-
- Pickart CM. Mechanisms underlying ubiquitination. Annual Review of Biochemistry. 2001;2001(70):195–201. - PubMed
-
- Christensen DE, Brzovic PS, Klevit RE. E2-BRCA1 RING interactions dictate synthesis of mono- or specific polyubiquitin chain linkages. Nature Structural and Molecular Biology. 2007;14:941–948. - PubMed
-
- Alpi AF, Pace PE, Babu MM, Patel KJ. Mechanistic insight into site-restricted monoubiquitination of FANCD2 by Ube2t, FANCL, and FANCI. Molecular Cell. 2008;32:767–777. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
