

Knockout of mouse Cyp3a gene enhances synthesis of cholesterol and bile acid in the liver
- PMID: 23709690
- PMCID: PMC3708357
- DOI: 10.1194/jlr.M033464
Knockout of mouse Cyp3a gene enhances synthesis of cholesterol and bile acid in the liver
Abstract
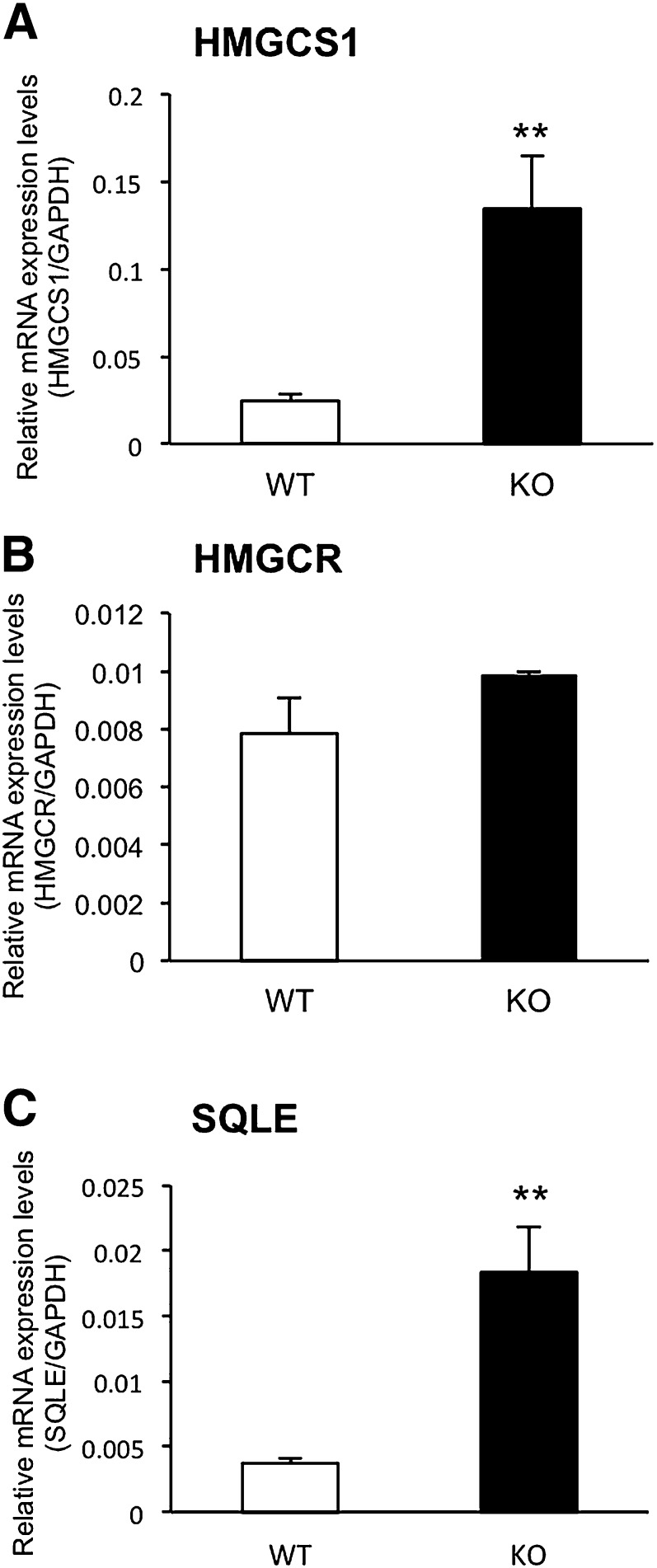
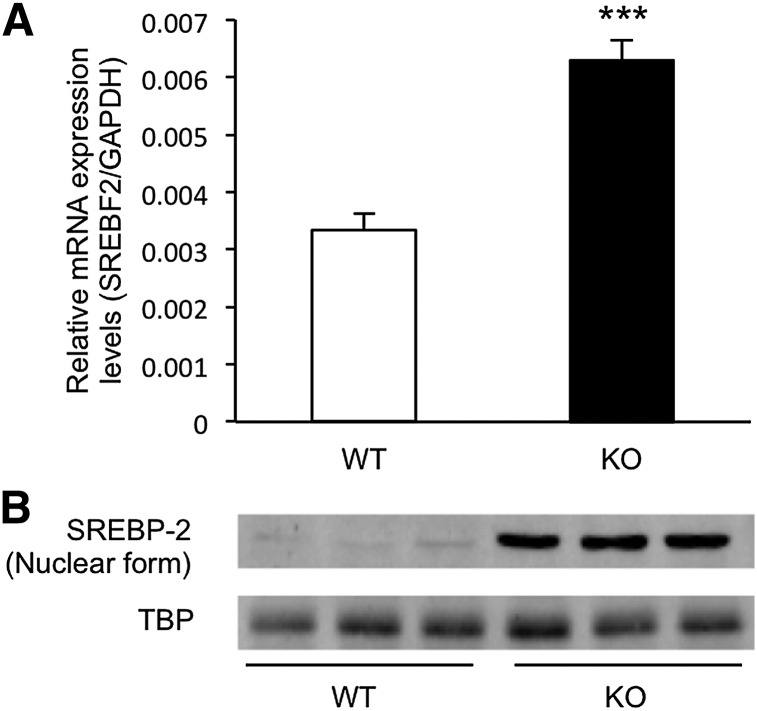
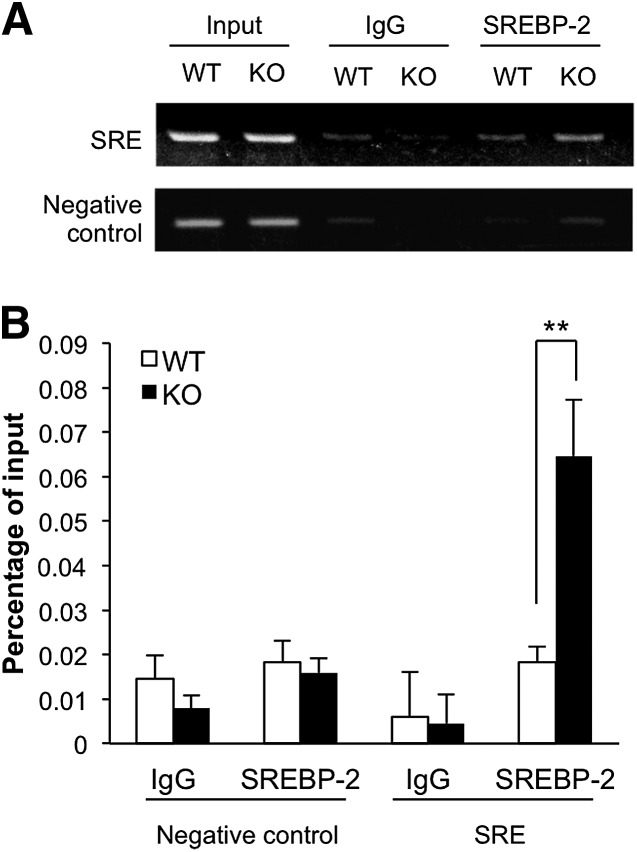
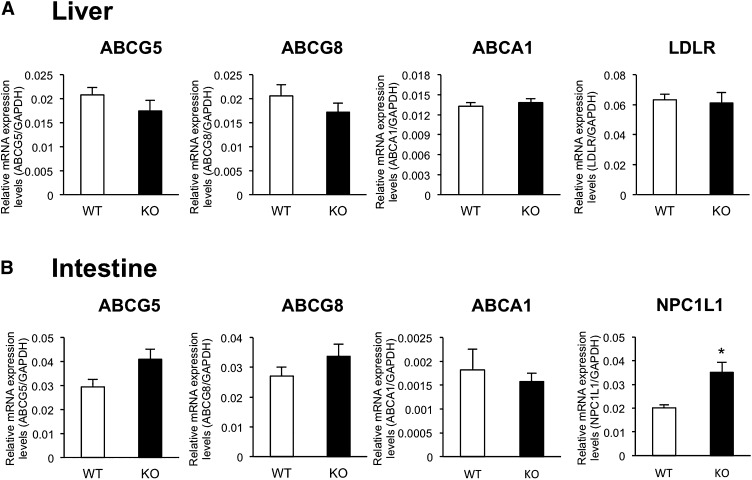
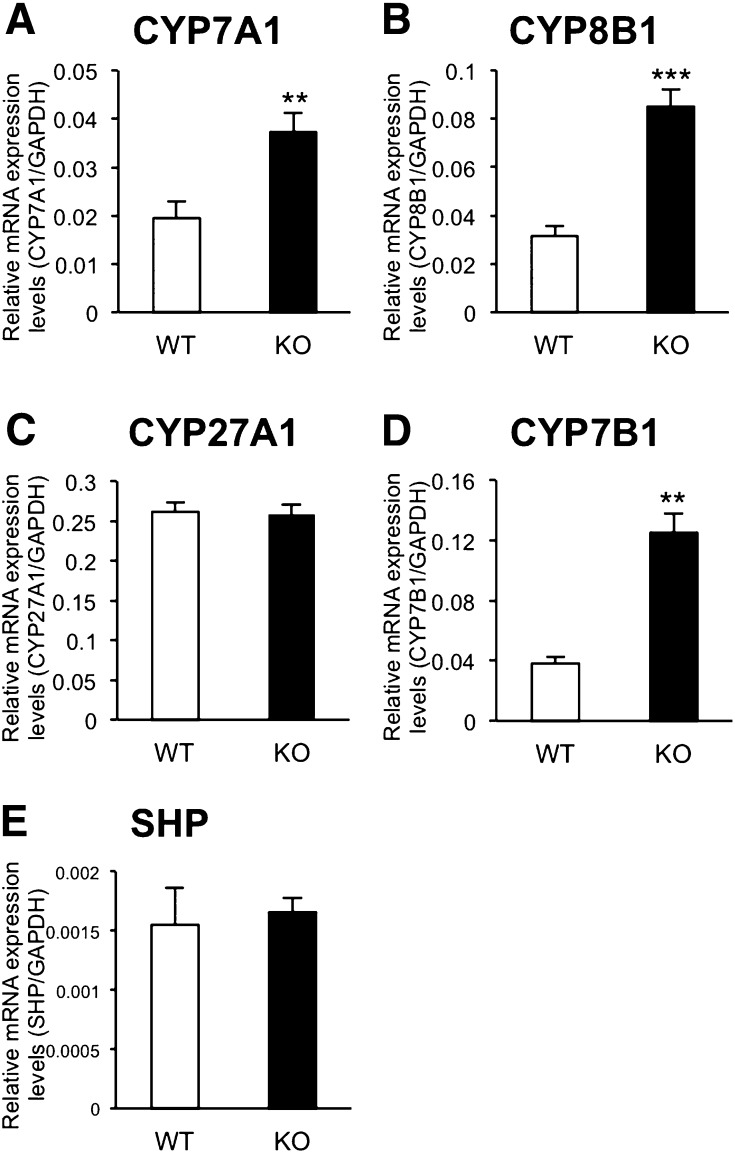
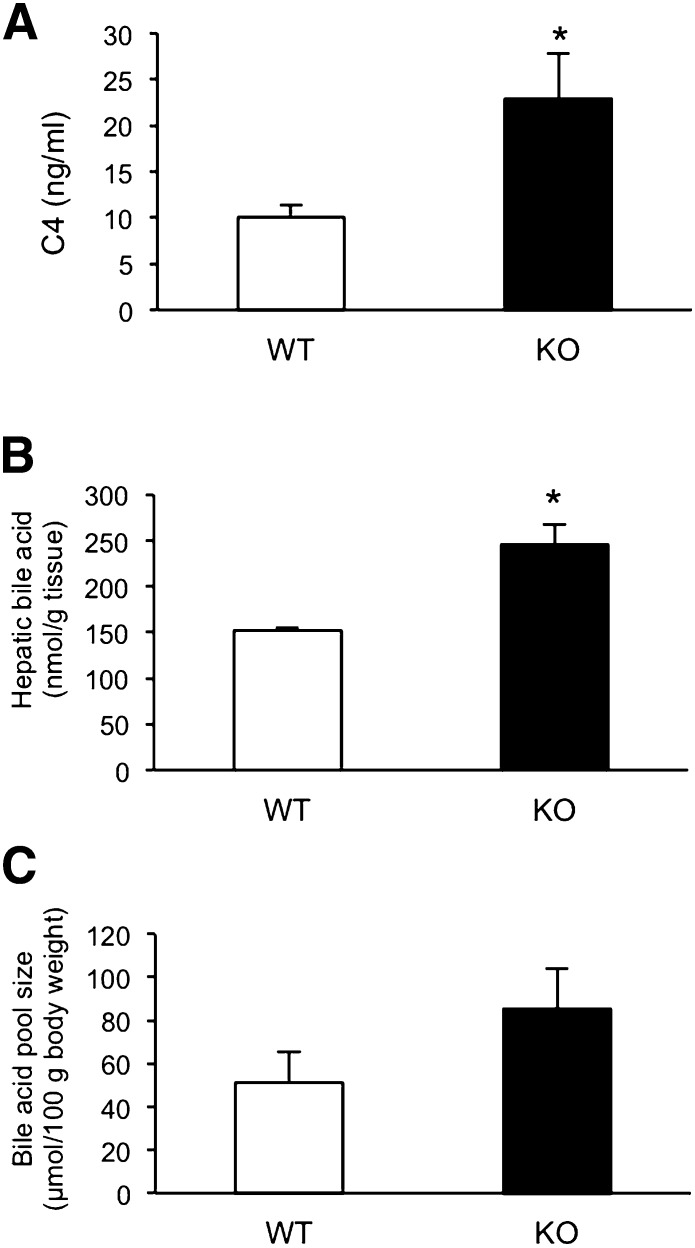
Here, we studied the effects of cytochrome P450 (CYP)3A deficiency on the mRNA expression of genes encoding regulators of hepatic cholesterol levels using Cyp3a-knockout (Cyp3a(-/-)) mice. The mRNA expression levels of genes encoding enzymes involved in cholesterol biosynthesis in the livers of Cyp3a(-/-) mice were higher than those of wild-type (WT) mice. Nuclear levels of sterol regulatory element-binding protein-2 (SREBP-2), which enhances cholesterol biosynthesis, were also higher in the livers of Cyp3a(-/-) mice. Binding of SREBP-2 to the Hmgcs1 gene promoter was more abundant in the livers of Cyp3a(-/-) mice. These results suggest that deficiency of CYP3A enzymes enhances transcription of genes encoding enzymes involved in cholesterol biosynthesis via activation of SREBP-2. On the other hand, hepatic cholesterol levels in Cyp3a(-/-) mice were 20% lower than those in WT mice. The mRNA expression levels of genes encoding enzymes involved in bile acid synthesis, plasma levels of 7α-hydroxy-4-cholesten-3-one and hepatic levels of total bile acid were significantly higher in Cyp3a(-/-) mice than in WT mice. These findings suggest that reduction of hepatic total cholesterol in Cyp3a(-/-) mice would be the consequence of enhanced bile acid synthesis. Therefore, CYP3A enzymes appear to play roles in the synthesis of cholesterol and bile acid in vivo.
Keywords: 25-hydroxy-cholesterol; HMG-CoA synthase 1; cytochrome P450 3A; squalene epoxidase; sterol regulatory element-binding protein-2.
Figures
References
-
- Miller W. L. 1988. Molecular biology of steroid hormone synthesis. Endocr. Rev. 9: 295–318 - PubMed
-
- Schroepfer G. J. 2000. Oxysterols: modulators of cholesterol metabolism and other processes. Physiol. Rev. 80: 361–554 - PubMed
-
- Bloch K., Berg B. N., Rittenberg D. 1943. The biological conversion of cholesterol to cholic acid. J. Biol. Chem. 149: 511–517
-
- Wiseman H. 1993. Vitamin D is a membrane antioxidant. Ability to inhibit iron-dependent lipid peroxidation in liposomes compared to cholesterol, ergosterol and tamoxifen and relevance to anticancer action. FEBS Lett. 326: 285–288 - PubMed
-
- Brown M. S., Goldstein J. L. 1997. The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell. 89: 331–340 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
