

Differentiation of CD8 memory T cells depends on Foxo1
- PMID: 23712431
- PMCID: PMC3674697
- DOI: 10.1084/jem.20130392
Differentiation of CD8 memory T cells depends on Foxo1
Abstract
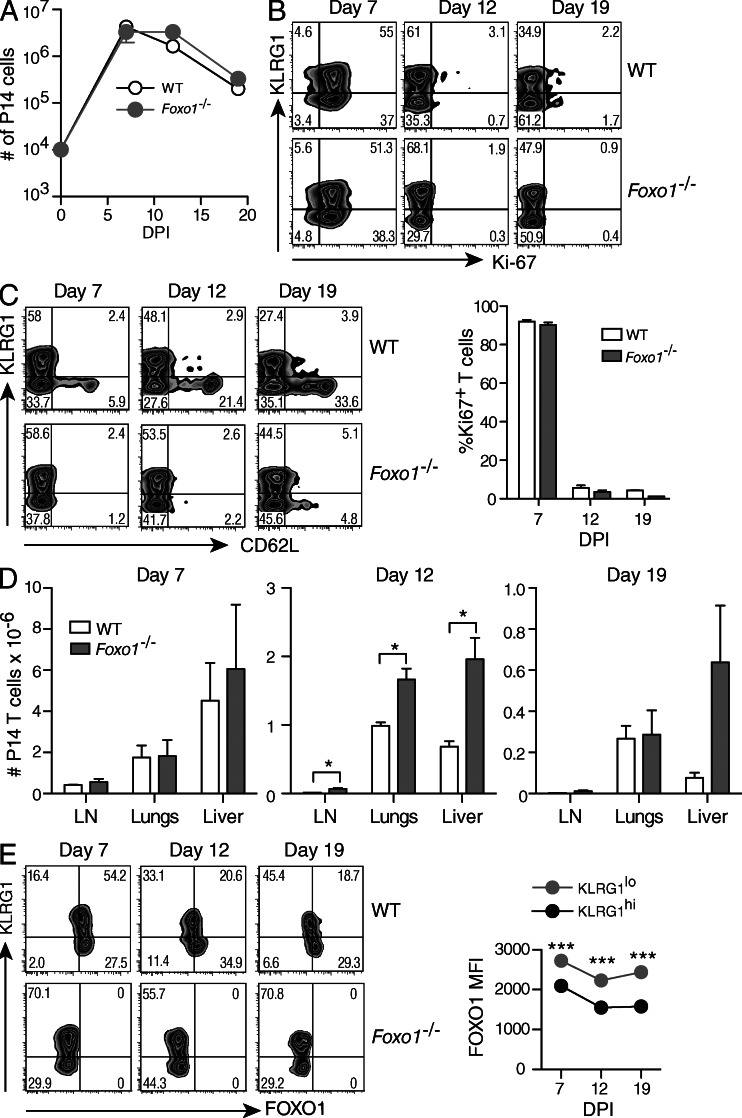
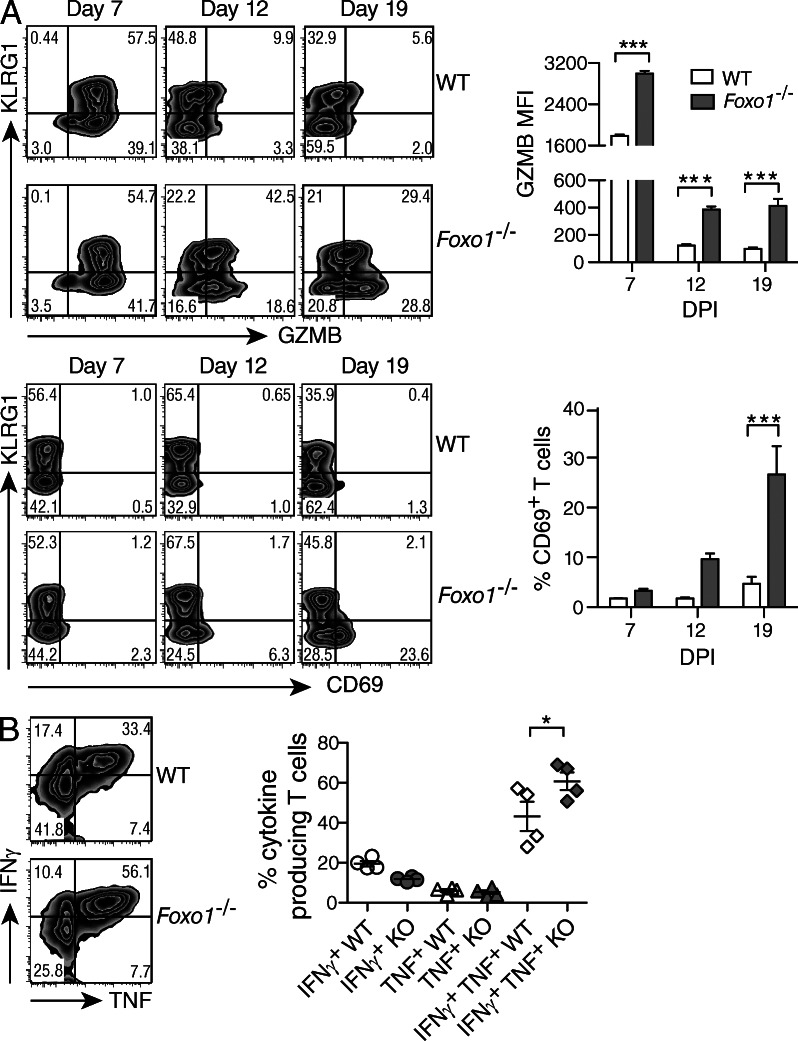
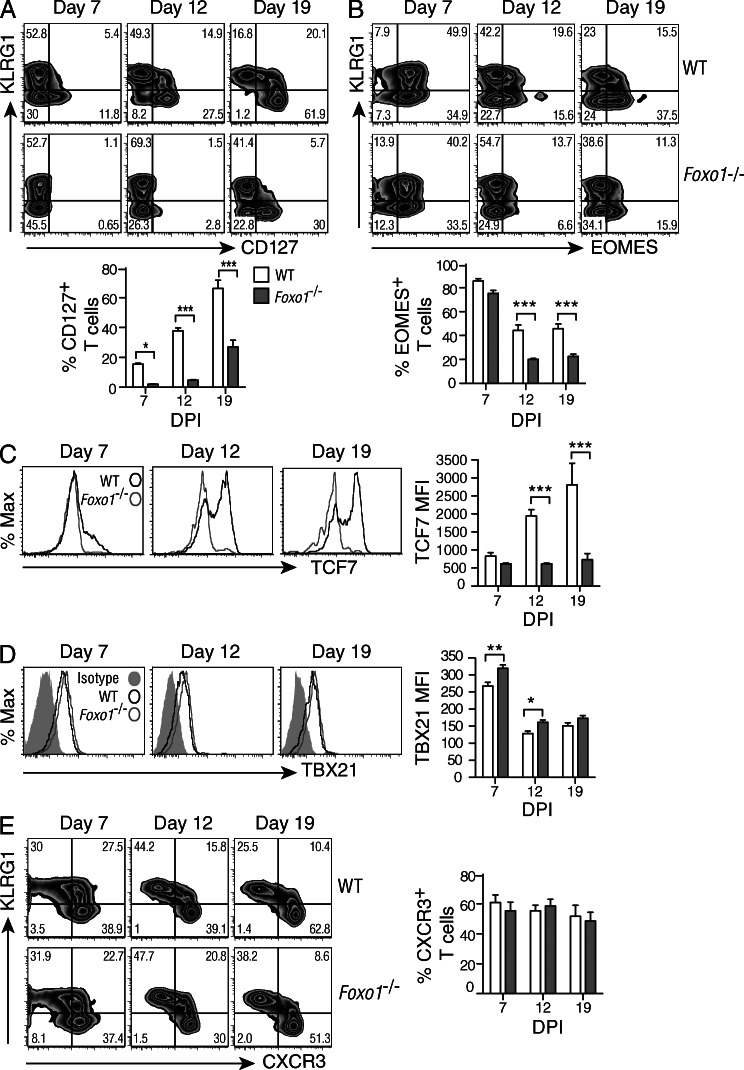
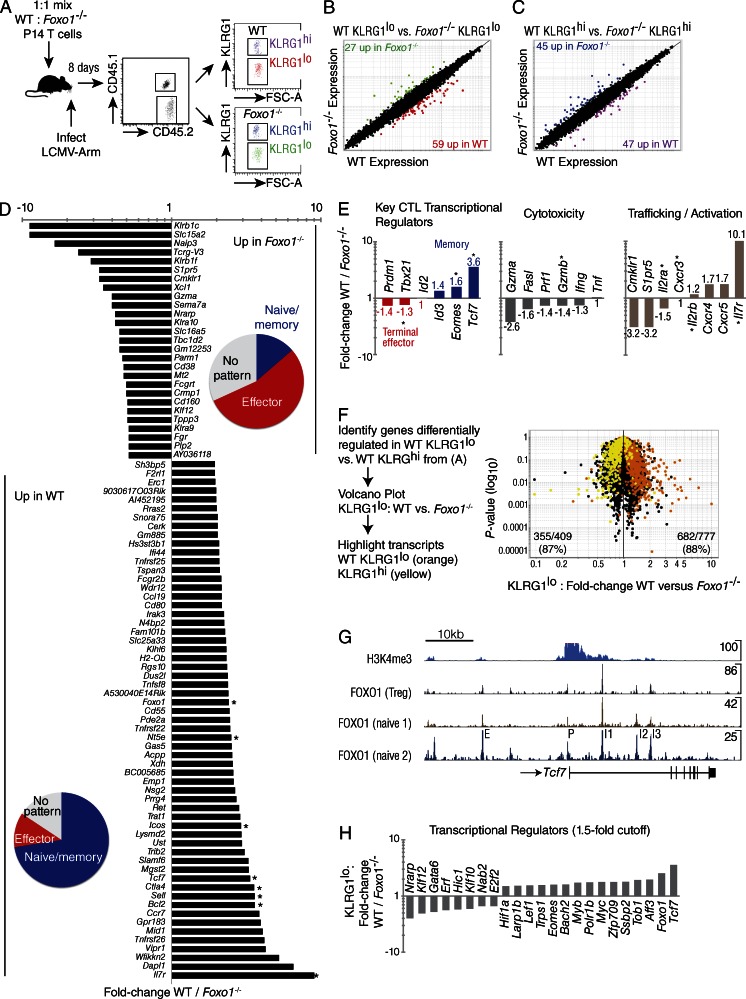
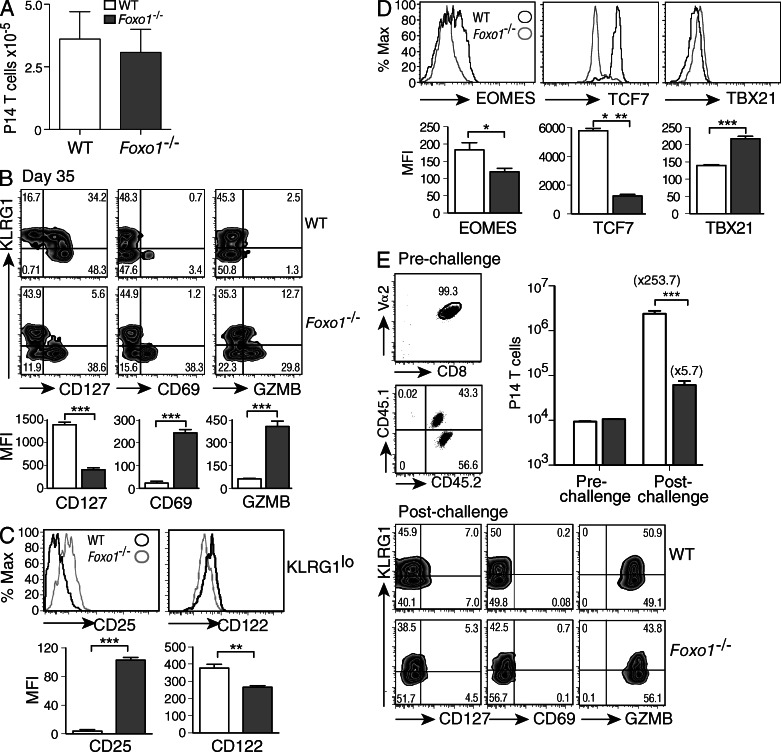
The forkhead O transcription factors (FOXO) integrate a range of extracellular signals, including growth factor signaling, inflammation, oxidative stress, and nutrient availability, to substantially alter the program of gene expression and modulate cell survival, cell cycle progression, and many yet to be unraveled cell type-specific responses. Naive antigen-specific CD8(+) T cells undergo a rapid expansion and arming of effector function within days of pathogen exposure. In addition, by the peak of expansion, they form precursors to memory T cells capable of self-renewal and indefinite survival. Using lymphocytic choriomeningitis virus Armstrong to probe the response to infection, we found that Foxo1(-/-) CD8(+) T cells expand normally with no defects in effector differentiation, but continue to exhibit characteristics of effector T cells long after antigen clearance. The KLRG1(lo) CD8(+) T cells that are normally enriched for memory-precursor cells retain Granzyme B and CD69 expression, and fail to up-regulate TCF7, EOMES, and other memory signature genes. As a correlate, Foxo1(-/-) CD8(+) T cells were virtually unable to expand upon secondary infection. Collectively, these results demonstrate an intrinsic role for FOXO1 in establishing the post-effector memory program that is essential to forming long-lived memory cells capable of immune reactivation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
