

Tdrkh is essential for spermatogenesis and participates in primary piRNA biogenesis in the germline
- PMID: 23714778
- PMCID: PMC3981179
- DOI: 10.1038/emboj.2013.121
Tdrkh is essential for spermatogenesis and participates in primary piRNA biogenesis in the germline
Abstract
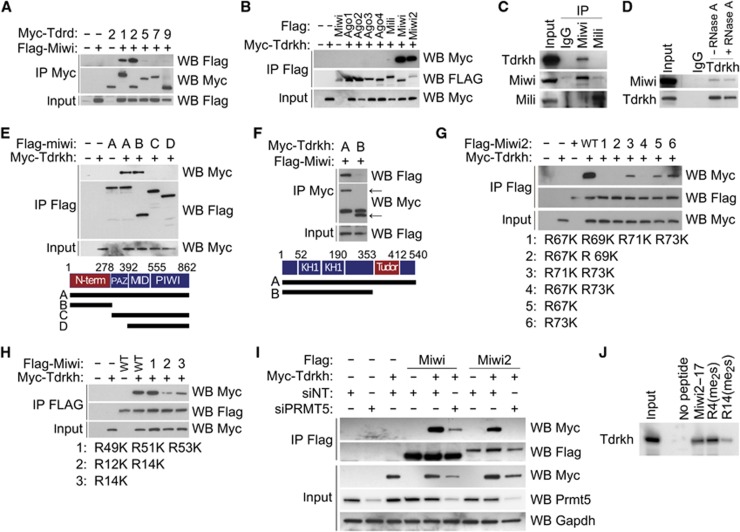
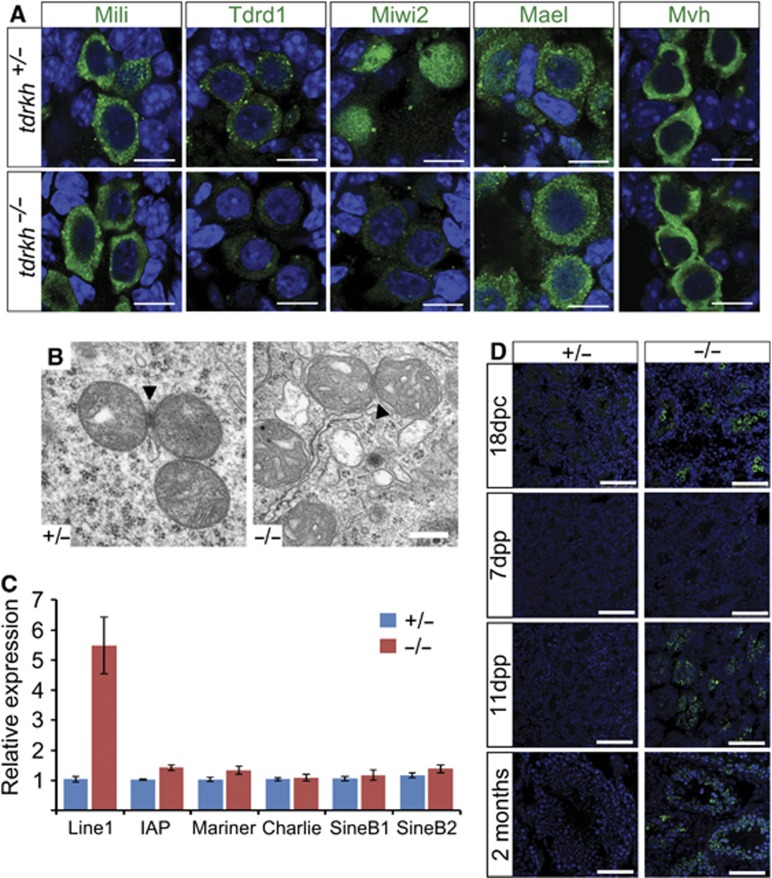
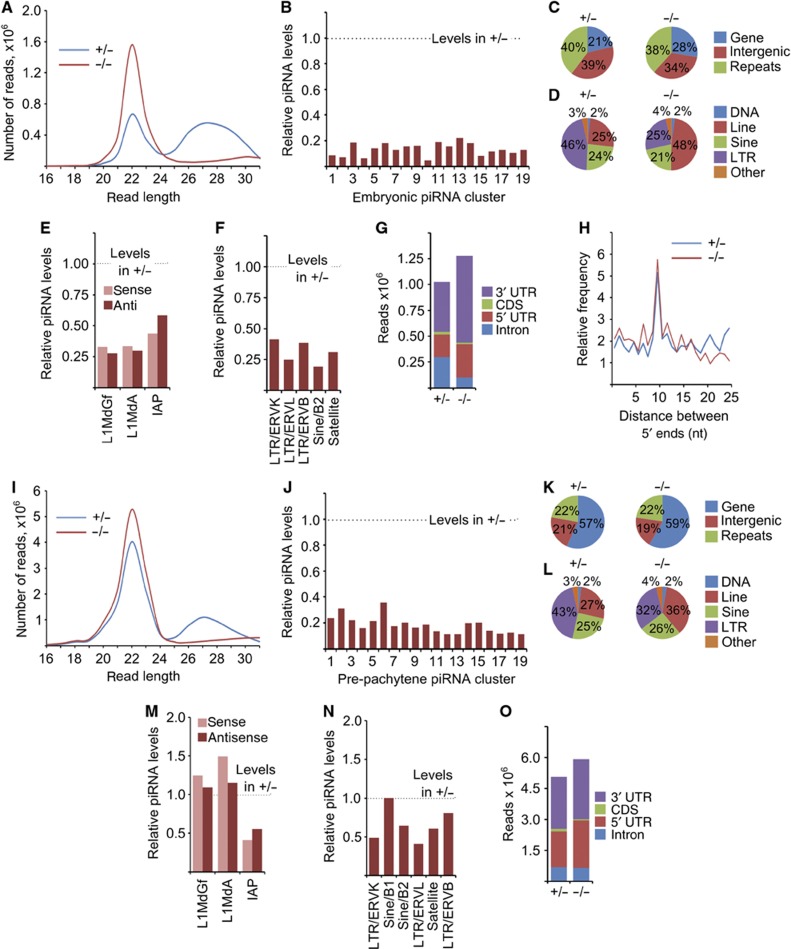
Piwi proteins and Piwi-interacting RNAs (piRNAs) repress transposition, regulate translation, and guide epigenetic programming in the germline. Here, we show that an evolutionarily conserved Tudor and KH domain-containing protein, Tdrkh (a.k.a. Tdrd2), is required for spermatogenesis and involved in piRNA biogenesis. Tdrkh partners with Miwi and Miwi2 via symmetrically dimethylated arginine residues in Miwi and Miwi2. Tdrkh is a mitochondrial protein often juxtaposed to pi-bodies and piP-bodies and is required for Tdrd1 cytoplasmic localization and Miwi2 nuclear localization. Tdrkh mutants display meiotic arrest at the zygotene stage, attenuate methylation of Line1 DNA, and upregulate Line1 RNA and protein, without inducing apoptosis. Furthermore, Tdrkh mutants have severely reduced levels of mature piRNAs but accumulate a distinct population of 1'U-containing, 2'O-methylated 31-37 nt RNAs that largely complement the missing mature piRNAs. Our results demonstrate that the primary piRNA biogenesis pathway involves 3'→5' processing of 31-37 nt intermediates and that Tdrkh promotes this final step of piRNA biogenesis but not the ping-pong cycle. These results shed light on mechanisms underlying primary piRNA biogenesis, an area in which information is conspicuously absent.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, Chien M, Russo JJ, Ju J, Sheridan R, Sander C, Zavolan M, Tuschl T (2006) A novel class of small RNAs bind to MILI protein in mouse testes. Nature 442: 203–207 - PubMed
-
- Aravin AA, Sachidanandam R, Girard A, Fejes-Toth K, Hannon GJ (2007) Developmentally regulated piRNA clusters implicate MILI in transposon control. Science 316: 744–747 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
