

How to make a functional β-cell
- PMID: 23715541
- PMCID: PMC3666377
- DOI: 10.1242/dev.093187
How to make a functional β-cell
Abstract
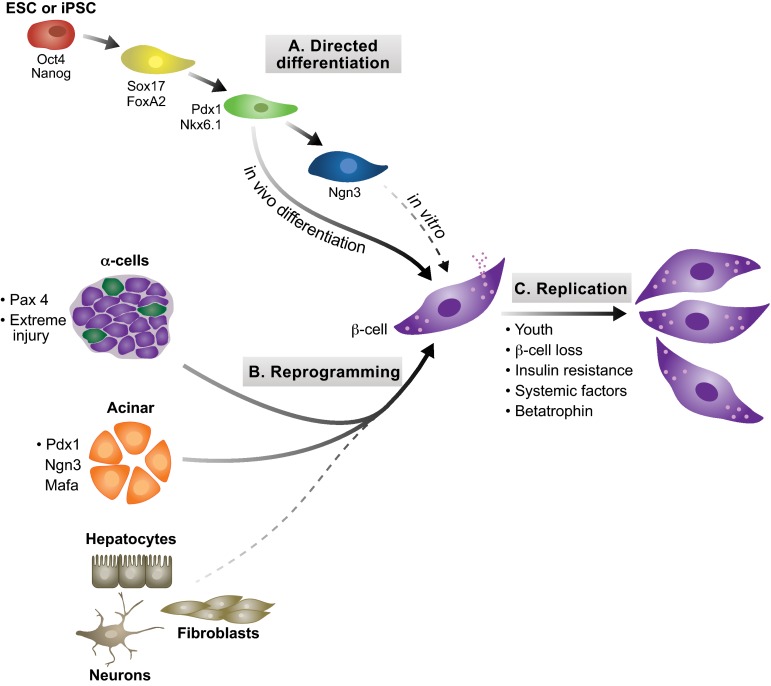
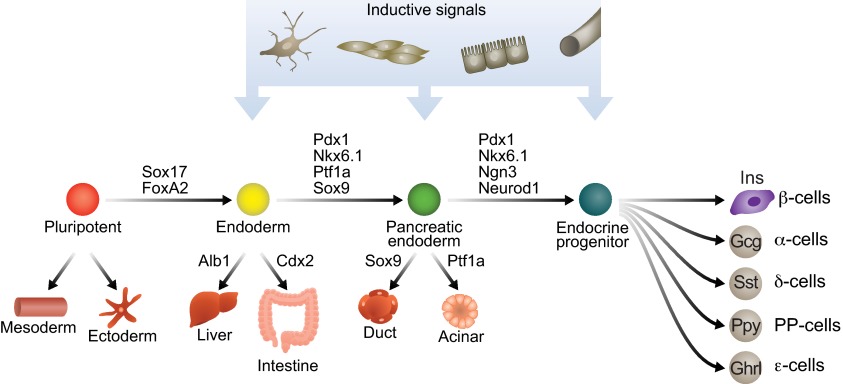
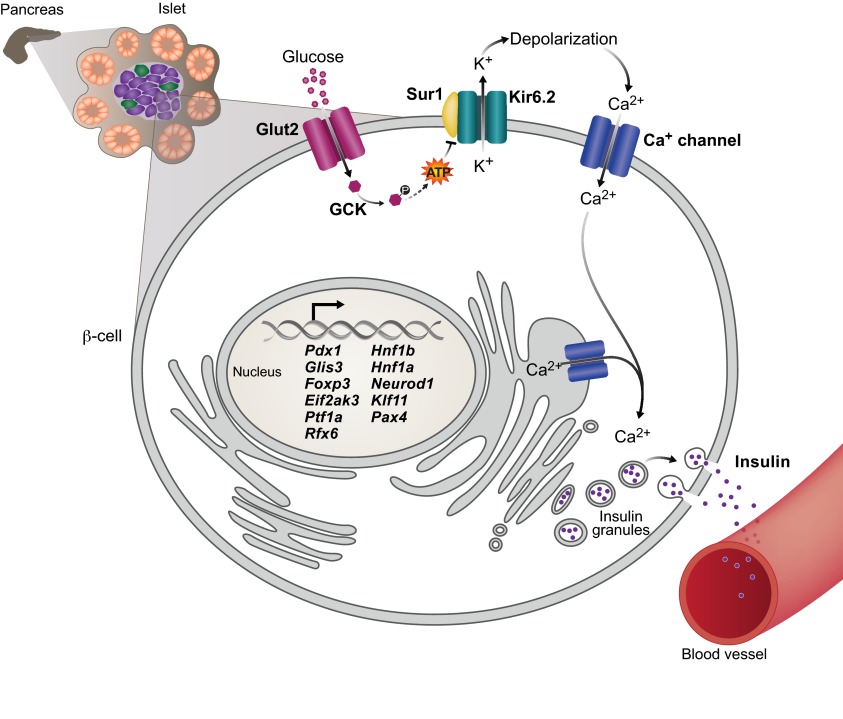
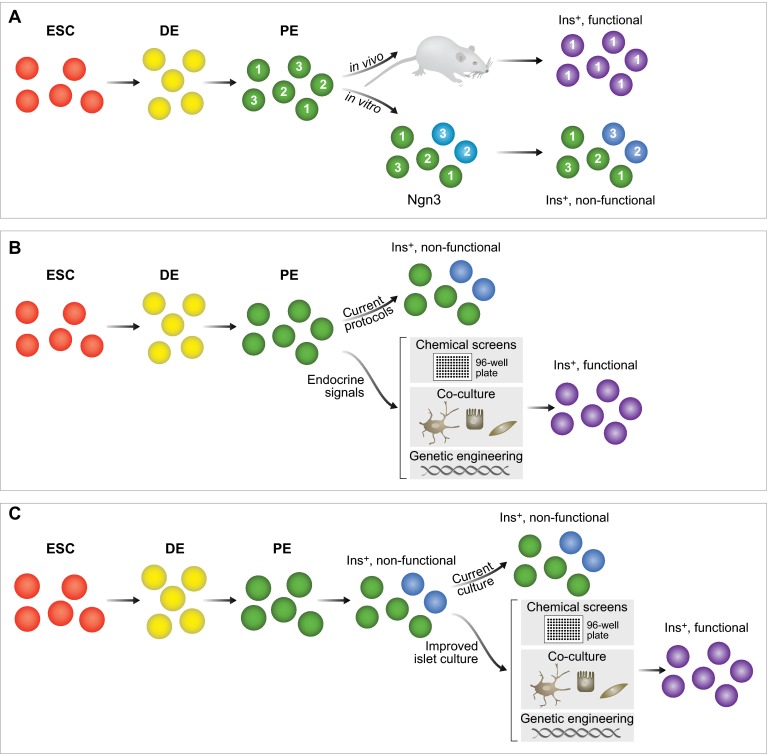
Insulin-secreting pancreatic β-cells are essential regulators of mammalian metabolism. The absence of functional β-cells leads to hyperglycemia and diabetes, making patients dependent on exogenously supplied insulin. Recent insights into β-cell development, combined with the discovery of pluripotent stem cells, have led to an unprecedented opportunity to generate new β-cells for transplantation therapy and drug screening. Progress has also been made in converting terminally differentiated cell types into β-cells using transcriptional regulators identified as key players in normal development, and in identifying conditions that induce β-cell replication in vivo and in vitro. Here, we summarize what is currently known about how these strategies could be utilized to generate new β-cells and highlight how further study into the mechanisms governing later stages of differentiation and the acquisition of functional capabilities could inform this effort.
Keywords: Diabetes mellitus; Mammalian metabolism; β cell.
Figures
References
-
- Ang S. L., Rossant J. (1994). HNF-3 beta is essential for node and notochord formation in mouse development. Cell 78, 561-574 - PubMed
-
- Apelqvist A., Ahlgren U., Edlund H. (1997). Sonic hedgehog directs specialised mesoderm differentiation in the intestine and pancreas. Curr. Biol. 7, 801-804 - PubMed
-
- Apelqvist A., Li H., Sommer L., Beatus P., Anderson D. J., Honjo T., Hrabe de Angelis M., Lendahl U., Edlund H. (1999). Notch signalling controls pancreatic cell differentiation. Nature 400, 877-881 - PubMed
-
- Araki R., Uda M., Hoki Y., Sunayama M., Nakamura M., Ando S., Sugiura M., Ideno H., Shimada A., Nifuji A., et al. (2013). Negligible immunogenicity of terminally differentiated cells derived from induced pluripotent or embryonic stem cells. Nature 494, 100-104 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
