

Mouse gene targeting reveals an essential role of mTOR in hematopoietic stem cell engraftment and hematopoiesis
- PMID: 23716557
- PMCID: PMC3762090
- DOI: 10.3324/haematol.2012.080424
Mouse gene targeting reveals an essential role of mTOR in hematopoietic stem cell engraftment and hematopoiesis
Abstract
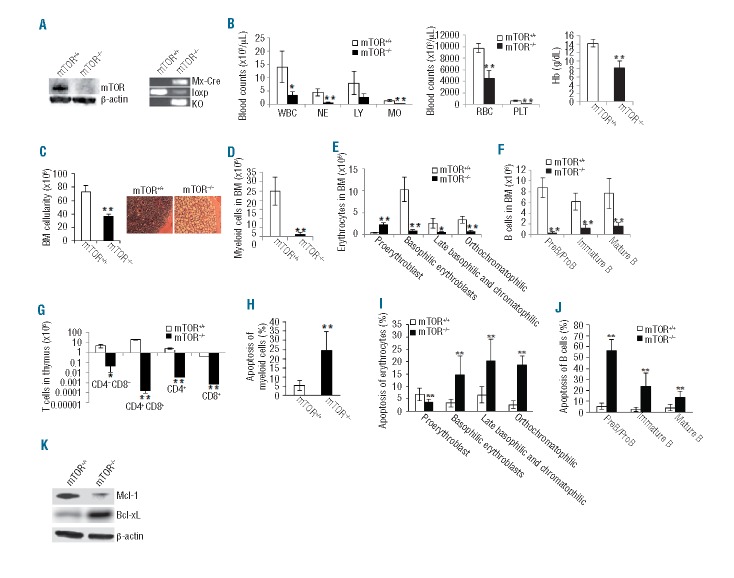
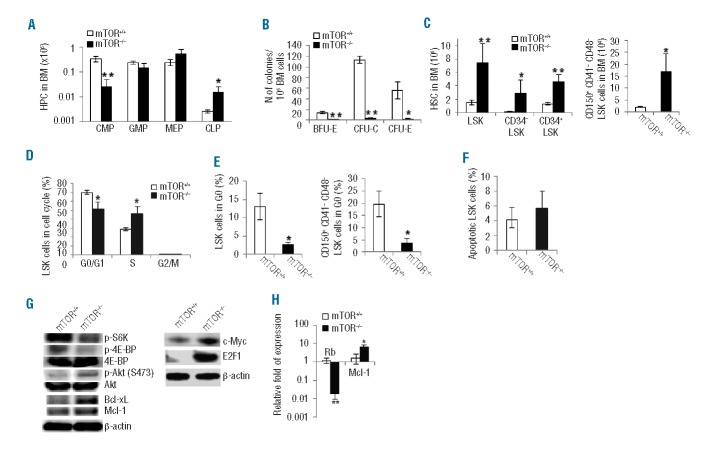
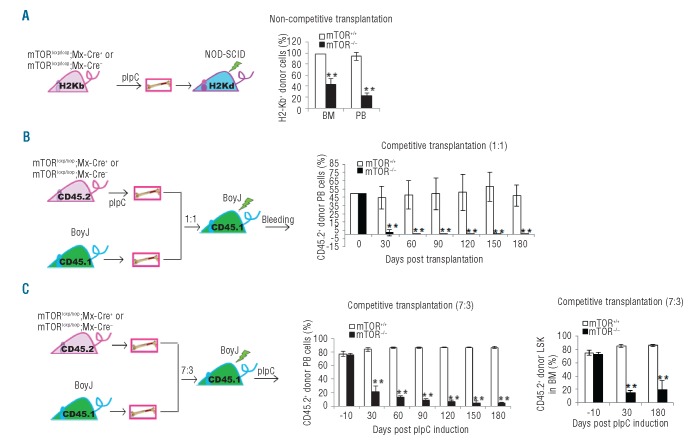
mTOR integrates signals from nutrients and growth factors to control protein synthesis, cell growth, and survival. Although mTOR has been established as a therapeutic target in hematologic malignancies, its physiological role in regulating hematopoiesis remains unclear. Here we show that conditional gene targeting of mTOR causes bone marrow failure and defects in multi-lineage hematopoiesis including myelopoiesis, erythropoiesis, thrombopoiesis, and lymphopoiesis. mTOR deficiency results in loss of quiescence of hematopoietic stem cells, leading to a transient increase but long-term exhaustion and defective engraftment of hematopoietic stem cells in lethally irradiated recipient mice. Furthermore, ablation of mTOR causes increased apoptosis in lineage-committed blood cells but not hematopoietic stem cells, indicating a differentiation stage-specific function. These results demonstrate that mTOR is essential for hematopoietic stem cell engraftment and multi-lineage hematopoiesis.
Figures
References
-
- Proud CG. The multifaceted role of mTOR in cellular stress responses. DNA Repair (Amst). 2004;3(8–9):927–34 - PubMed
-
- Raslova H, Baccini V, Loussaief L, Comba B, Larghero J, Debili N, et al. Mammalian target of rapamycin (mTOR) regulates both proliferation of megakaryocyte progenitors and late stages of megakaryocyte differentiation. Blood. 2006;107(6):2303–10 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
