

Congenital erythrocytosis associated with gain-of-function HIF2A gene mutations and erythropoietin levels in the normal range
- PMID: 23716564
- PMCID: PMC3789469
- DOI: 10.3324/haematol.2013.088369
Congenital erythrocytosis associated with gain-of-function HIF2A gene mutations and erythropoietin levels in the normal range
Abstract
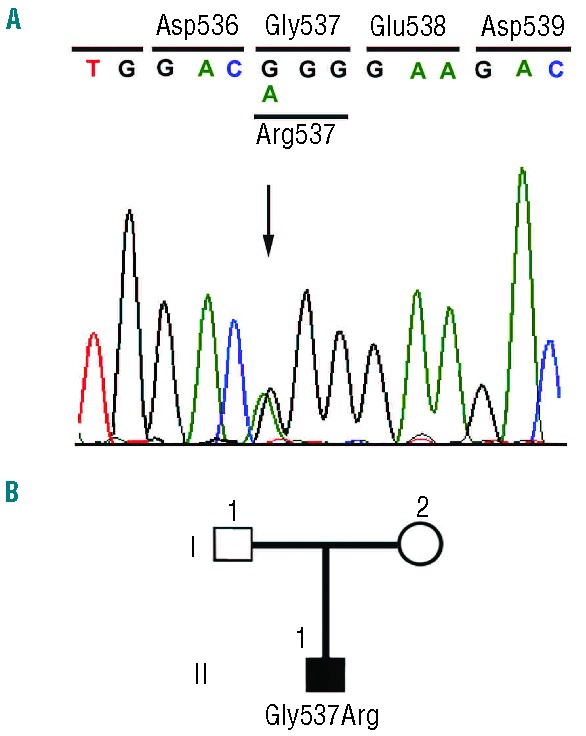
Hypoxia-inducible factor 2α (HIF-2α) plays a pivotal role in the balancing of oxygen requirements throughout the body. The protein is a transcription factor that modulates the expression of a wide array of genes and, in turn, controls several key processes including energy metabolism, erythropoiesis and angiogenesis. We describe here the identification of two cases of familial erythrocytosis associated with heterozygous HIF2A missense mutations, namely Ile533Val and Gly537Arg. Ile533Val is a novel mutation and represents the genetic HIF2A change nearest to Pro-531, the primary hydroxyl acceptor residue, so far identified. The Gly537Arg missense mutation has already been described in familial erythrocytosis. However, our patient is the only described case of a de novo HIF2A mutation associated with the development of congenital polycythemia. Functional in vivo studies, based on exogenous expression of hybrid HIF-2α transcription factors, indicated that these genetic alterations lead to the stabilization of HIF-2α protein. All the identified polycythemic subjects with HIF2A mutations show serum erythropoietin in the normal range, independently of the hematocrit values and phlebotomy frequency. The erythroid precursors obtained from the peripheral blood of patients showed an altered phenotype, including an increased rate of growth and a modified expression of some HIF-2α target genes. These results suggest the novel proposal that polycythemia observed in subjects with HIF2A mutations might also be due to primary changes in hematopoietic cells and not only secondary to increased erythropoietin levels.
Figures





References
-
- Abbrecht PH, Littell JK. Plasma erythropoietin in men and mice during acclimatization to different altitudes. J Appl Physiol. 1972;32(1):54–8 - PubMed
-
- Jelkmann W. Erythropoietin after a century of research: younger than ever. Eur J Haematol. 2007;78(3):183–205 - PubMed
-
- Maxwell PH, Osmond MK, Pugh CW, Heryet A, Nicholls LG, Tan CC, et al. Identification of the renal erythropoietin-producing cells using transgenic mice. Kidney Int. 1993;44(5):1149–62 - PubMed
-
- Wenger RH, Hoogewijs D. Regulated oxygen sensing by protein hydroxylation in renal erythropoietin-producing cells. Am J Physiol Renal Physiol. 2010; 298(6):F1287–96 - PubMed
-
- Obara N, Suzuki N, Kim K, Nagasawa T, Imagawa S, Yamamoto M. Repression via the GATA box is essential for tissue-specific erythropoietin gene expression. Blood. 2008;111(10):5223–32 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
