

Receptor-like kinase SOBIR1/EVR interacts with receptor-like proteins in plant immunity against fungal infection
- PMID: 23716655
- PMCID: PMC3683720
- DOI: 10.1073/pnas.1220015110
Receptor-like kinase SOBIR1/EVR interacts with receptor-like proteins in plant immunity against fungal infection
Erratum in
- Proc Natl Acad Sci U S A. 2013 Aug 6;110(32):13228. America, Antione H P [corrected to America, Antoine H P]
Abstract
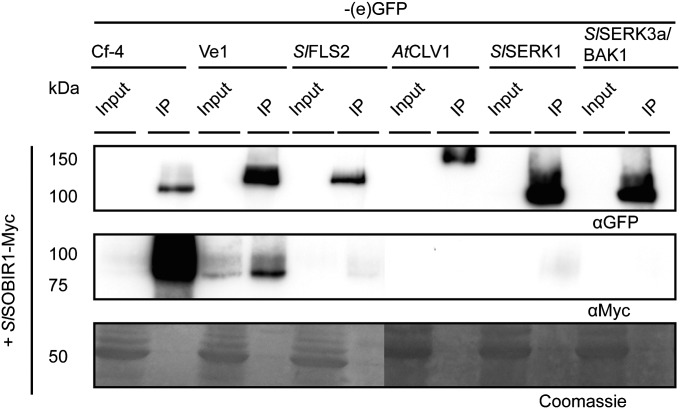
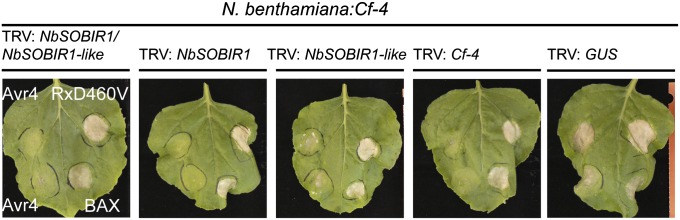
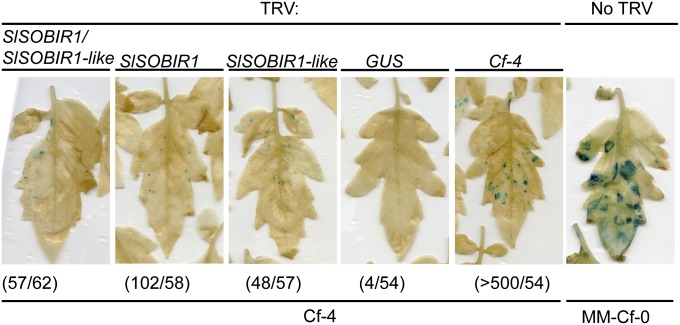
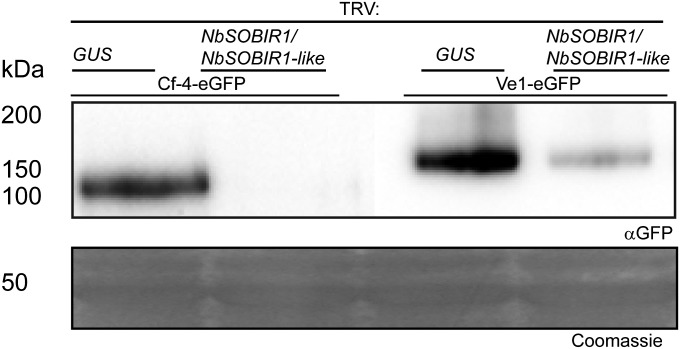
The plant immune system is activated by microbial patterns that are detected as nonself molecules. Such patterns are recognized by immune receptors that are cytoplasmic or localized at the plasma membrane. Cell surface receptors are represented by receptor-like kinases (RLKs) that frequently contain extracellular leucine-rich repeats and an intracellular kinase domain for activation of downstream signaling, as well as receptor-like proteins (RLPs) that lack this signaling domain. It is therefore hypothesized that RLKs are required for RLPs to activate downstream signaling. The RLPs Cf-4 and Ve1 of tomato (Solanum lycopersicum) mediate resistance to the fungal pathogens Cladosporium fulvum and Verticillium dahliae, respectively. Despite their importance, the mechanism by which these immune receptors mediate downstream signaling upon recognition of their matching ligand, Avr4 and Ave1, remained enigmatic. Here we show that the tomato ortholog of the Arabidopsis thaliana RLK Suppressor Of BIR1-1/Evershed (SOBIR1/EVR) and its close homolog S. lycopersicum (Sl)SOBIR1-like interact in planta with both Cf-4 and Ve1 and are required for the Cf-4- and Ve1-mediated hypersensitive response and immunity. Tomato SOBIR1/EVR interacts with most of the tested RLPs, but not with the RLKs FLS2, SERK1, SERK3a, BAK1, and CLV1. SOBIR1/EVR is required for stability of the Cf-4 and Ve1 receptors, supporting our observation that these RLPs are present in a complex with SOBIR1/EVR in planta. We show that SOBIR1/EVR is essential for RLP-mediated immunity and propose that the protein functions as a regulatory RLK of this type of cell-surface receptors.
Keywords: defense signaling; development; plant innate immunity; plant–microbe interaction; receptor complex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Jones JDG, Dangl JL. The plant immune system. Nature. 2006;444(7117):323–329. - PubMed
-
- Boller T, Felix G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu Rev Plant Biol. 2009;60:379–406. - PubMed
-
- Göhre V, Robatzek S. Breaking the barriers: Microbial effector molecules subvert plant immunity. Annu Rev Phytopathol. 2008;46:189–215. - PubMed
-
- de Jonge R, Bolton MD, Thomma BPHJ. How filamentous pathogens co-opt plants: The ins and outs of fungal effectors. Curr Opin Plant Biol. 2011;14(4):400–406. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
