

Metabolic aspects of neuron-oligodendrocyte-astrocyte interactions
- PMID: 23717302
- PMCID: PMC3651962
- DOI: 10.3389/fendo.2013.00054
Metabolic aspects of neuron-oligodendrocyte-astrocyte interactions
Abstract
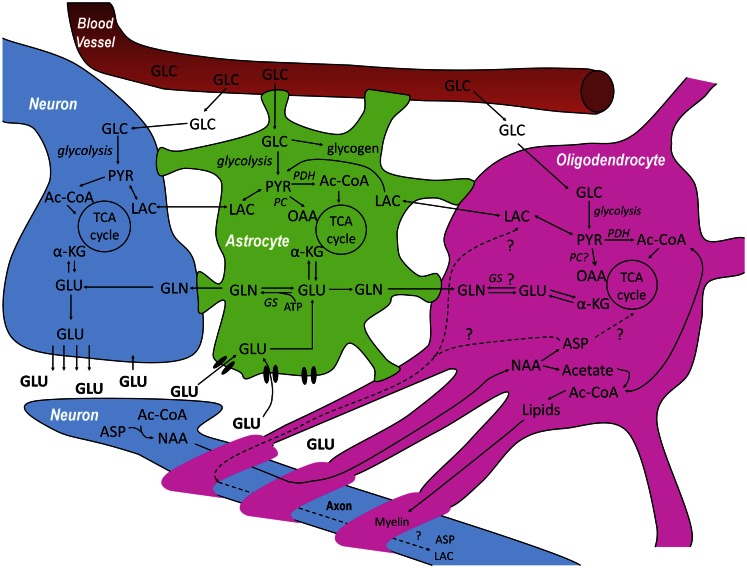
Whereas astrocytes have been in the limelight of scientific interest in brain energy metabolism for a while, oligodendrocytes are still waiting for a place on the metabolic stage. We propose to term the interaction of oligodendrocytes with astrocytes and neurons: NOA (neuron-oligodendrocyte-astrocyte) interactions. One of the reasons to find out more about metabolic interactions between oligodendrocytes, neurons, and astrocytes is to establish markers of healthy oligodendrocyte metabolism that could be used for the diagnosis and assessment of white matter disease. The vesicular release of glutamate in the white matter has received considerable attention in the past. Oligodendrocyte lineage cells express glutamate receptors and glutamate toxicity has been implicated in diseases affecting oligodendrocytes such as hypoxic-ischaemic encephalopathy, inflammatory diseases and trauma. As oligodendrocyte precursor cells vividly react to injury it is also important to establish whether cells recruited into damaged areas are able to regenerate lost myelin sheaths or whether astrocytic scarring occurs. It is therefore important to consider metabolic aspects of astrocytes and oligodendrocytes separately. The present review summarizes the limited evidence available on metabolic cycles in oligodendrocytes and so hopes to stimulate further research interests in this important field.
Keywords: energy metabolism; glucose; glycolysis; gray matter; lactate; monocarboxylate transporters; pyruvate carboxylation; white matter.
Figures
References
-
- Brekke E. M., Walls A. B., Schousboe A., Waagepetersen H. S., Sonnewald U. (2012). Quantitative importance of the pentose phosphate pathway determined by incorporation of 13C from [2-13C]- and [3-13C]glucose into TCA cycle intermediates and neurotransmitter amino acids in functionally intact neurons. J. Cereb. Blood Flow Metab. 32, 1788–1799 10.1038/jcbfm.2012.85 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous