

When stress and development go hand in hand: main hormonal controls of adventitious rooting in cuttings
- PMID: 23717317
- PMCID: PMC3653114
- DOI: 10.3389/fpls.2013.00133
When stress and development go hand in hand: main hormonal controls of adventitious rooting in cuttings
Abstract
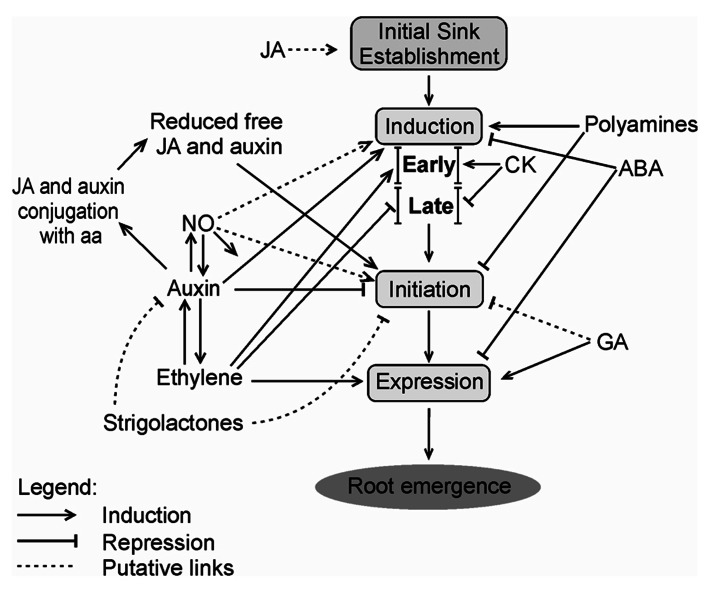
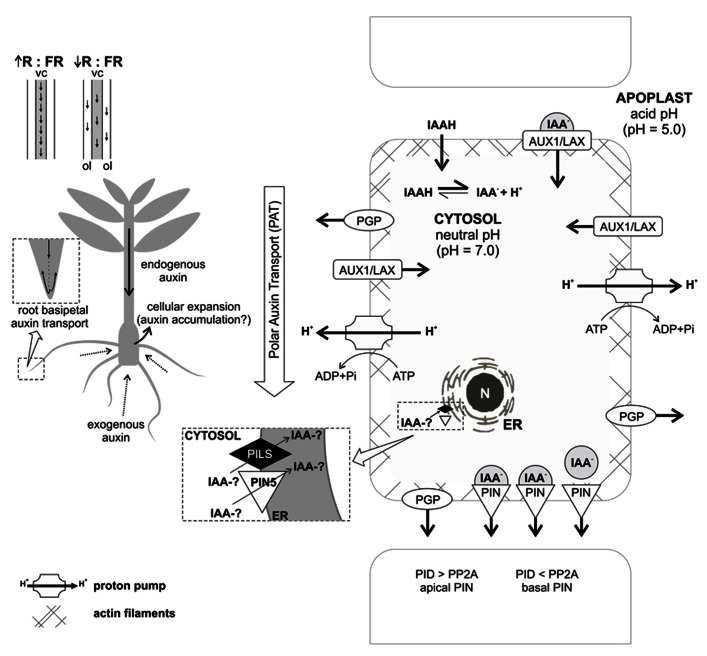
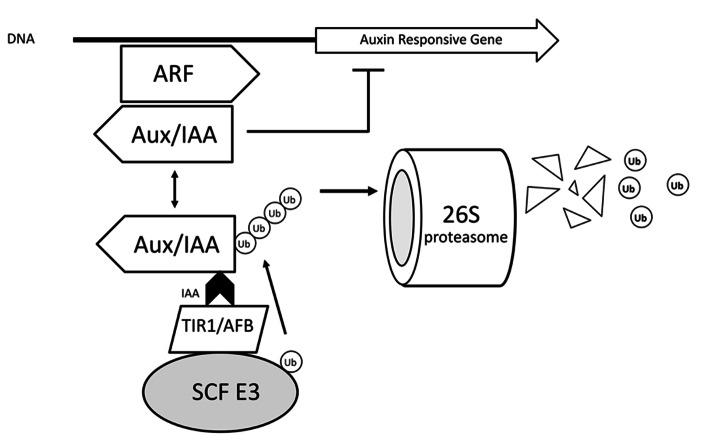
Adventitious rooting (AR) is a multifactorial response leading to new roots at the base of stem cuttings, and the establishment of a complete and autonomous plant. AR has two main phases: (a) induction, with a requirement for higher auxin concentration; (b) formation, inhibited by high auxin and in which anatomical changes take place. The first stages of this process in severed organs necessarily include wounding and water stress responses which may trigger hormonal changes that contribute to reprogram target cells that are competent to respond to rooting stimuli. At severance, the roles of jasmonate and abscisic acid are critical for wound response and perhaps sink strength establishment, although their negative roles on the cell cycle may inhibit root induction. Strigolactones may also inhibit AR. A reduced concentration of cytokinins in cuttings results from the separation of the root system, whose tips are a relevant source of these root induction inhibitors. The combined increased accumulation of basipetally transported auxins from the shoot apex at the cutting base is often sufficient for AR in easy-to-root species. The role of peroxidases and phenolic compounds in auxin catabolism may be critical at these early stages right after wounding. The events leading to AR strongly depend on mother plant nutritional status, both in terms of minerals and carbohydrates, as well as on sink establishment at cutting bases. Auxins play a central role in AR. Auxin transporters control auxin canalization to target cells. There, auxins act primarily through selective proteolysis and cell wall loosening, via their receptor proteins TIR1 (transport inhibitor response 1) and ABP1 (Auxin-Binding Protein 1). A complex microRNA circuitry is involved in the control of auxin response factors essential for gene expression in AR. After root establishment, new hormonal controls take place, with auxins being required at lower concentrations for root meristem maintenance and cytokinins needed for root tissue differentiation.
Keywords: adventitious rooting; auxin; cytokinin; hormonal crosstalk; jasmonic acid; microRNAs; nutrition; receptors.
Figures

References
-
- Abu-Abied M., Szwerdszarf D., Mordehaev I., Levy A., Rogovoy O., Belausov E., et al. (2012). Microarray analysis revealed upregulation of nitrate reductase in juvenile cuttings of Eucalyptus grandis, which correlated with increased nitric oxide production and adventitious root formation. Plant J. 71 787–799 - PubMed
-
- Agulló-Antón, M. A., Sánchez-Bravo J., Acosta M., Druege U. (2011). Auxins or sugars: what makes the difference in the adventitious rooting of stored carnation cuttings? J. Plant Growth Regul. 30 100–113
-
- Ahkami A. H., Lischewski S., Haensch K. T., Porfirova S., Hofmann J., Rolletschek H., et al. (2009). Molecular physiology of adventitious root formation in Petunia hybrida cuttings: involvement of wound response and primary metabolism. New Phytol. 181 613–625 - PubMed
-
- Aida M., Beis D., Heidstra R., Willemsen V., Blilou I., Galinha C., et al. (2004). The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 119 109–120 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous