

Open or close the gate - stomata action under the control of phytohormones in drought stress conditions
- PMID: 23717320
- PMCID: PMC3652521
- DOI: 10.3389/fpls.2013.00138
Open or close the gate - stomata action under the control of phytohormones in drought stress conditions
Abstract
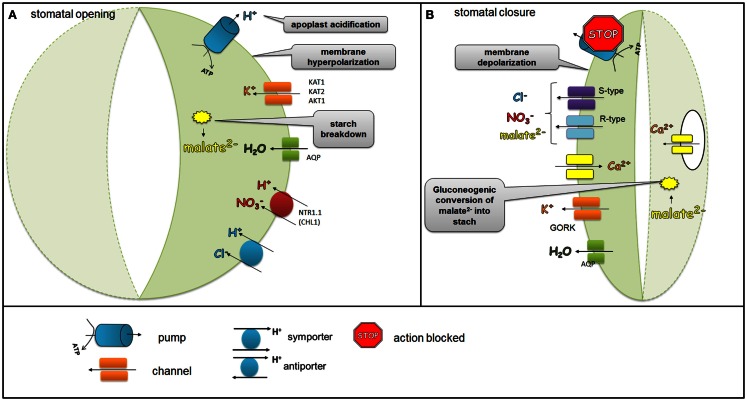
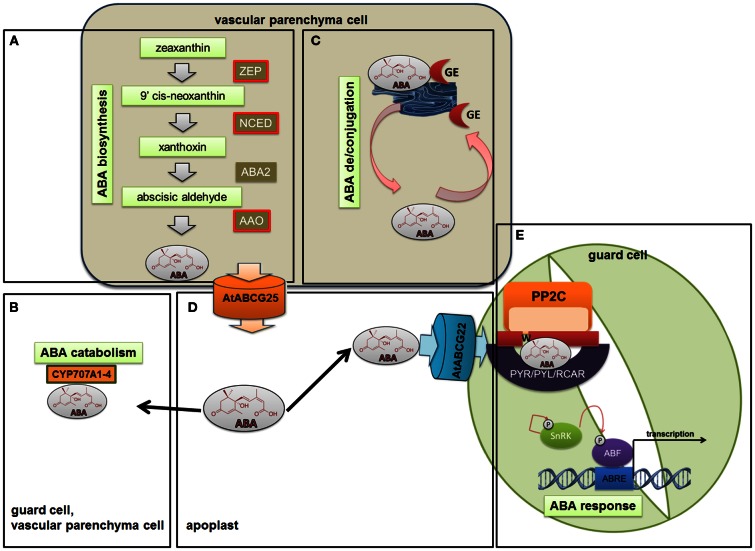
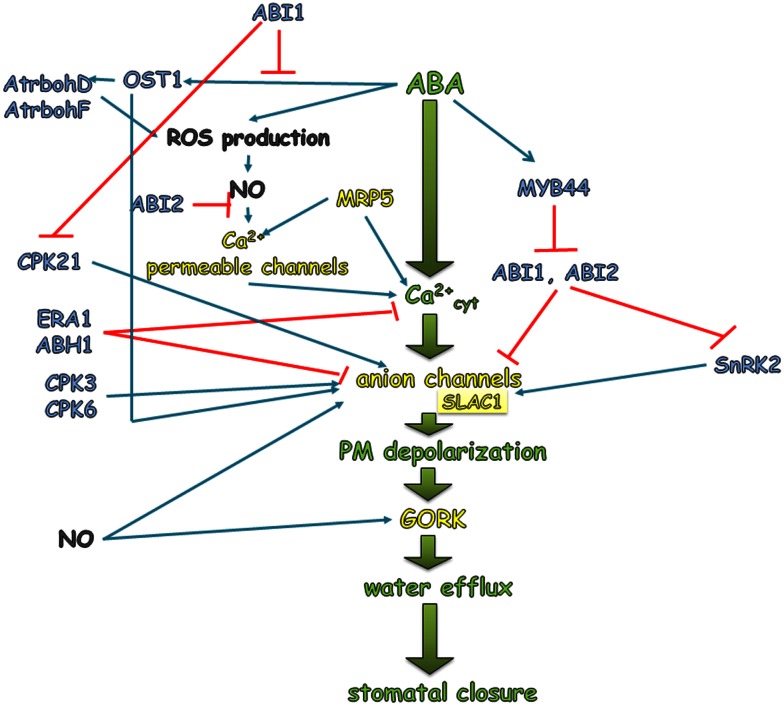
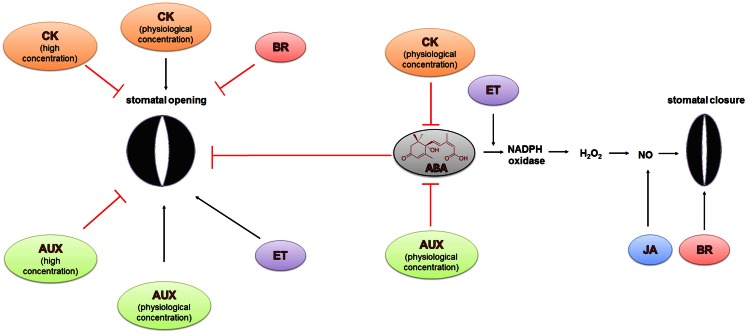
Two highly specialized cells, the guard cells that surround the stomatal pore, are able to integrate environmental and endogenous signals in order to control the stomatal aperture and thereby the gas exchange. The uptake of CO2 is associated with a loss of water by leaves. Control of the size of the stomatal aperture optimizes the efficiency of water use through dynamic changes in the turgor of the guard cells. The opening and closing of stomata is regulated by the integration of environmental signals and endogenous hormonal stimuli. The various different factors to which the guard cells respond translates into the complexity of the network of signaling pathways that control stomatal movements. The perception of an abiotic stress triggers the activation of signal transduction cascades that interact with or are activated by phytohormones. Among these, abscisic acid (ABA), is the best-known stress hormone that closes the stomata, although other phytohormones, such as jasmonic acid, brassinosteroids, cytokinins, or ethylene are also involved in the stomatal response to stresses. As a part of the drought response, ABA may interact with jasmonic acid and nitric oxide in order to stimulate stomatal closure. In addition, the regulation of gene expression in response to ABA involves genes that are related to ethylene, cytokinins, and auxin signaling. In this paper, recent findings on phytohormone crosstalk, changes in signaling pathways including the expression of specific genes and their impact on modulating stress response through the closing or opening of stomata, together with the highlights of gaps that need to be elucidated in the signaling network of stomatal regulation, are reviewed.
Keywords: ABA; abiotic stress; crosstalk; guard cells; jasmonic acid; phytohormones; stomata.
Figures


References
-
- Abeles F. B., Morgan P. W., Saltveit M. E. (1992). Ethylene in Plant Biology. 2nd Edn. San Diego: Academic Press
-
- Blatt M. R., Armstrong F. (1993). K+ channels of stomatal guard cells: abscisic-acid evoked control of the outward rectifier mediated by cytoplasmic pH. Planta 191, 330–341 10.1007/BF00195690 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials