

Glucose homeostasis in mice is transglutaminase 2 independent
- PMID: 23717413
- PMCID: PMC3661676
- DOI: 10.1371/journal.pone.0063346
Glucose homeostasis in mice is transglutaminase 2 independent
Abstract
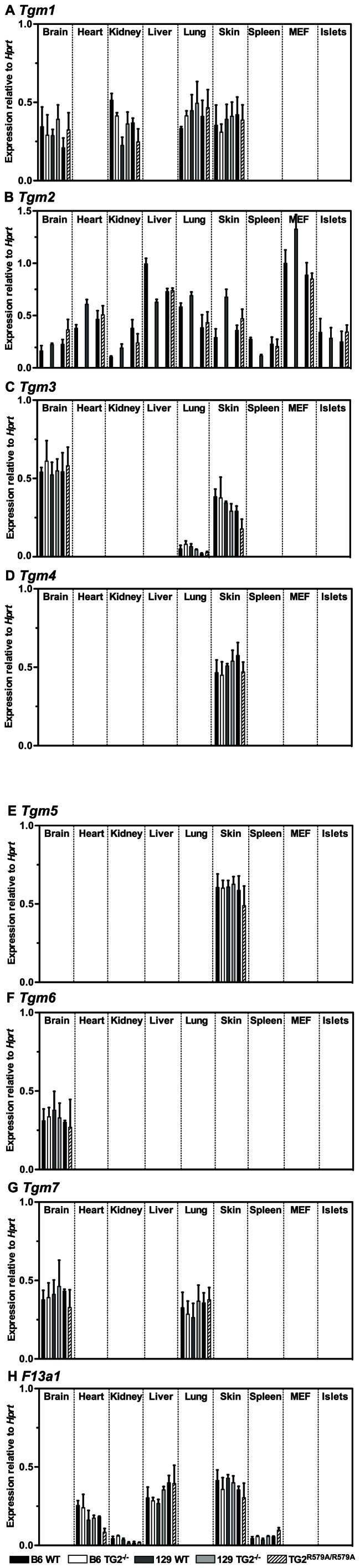
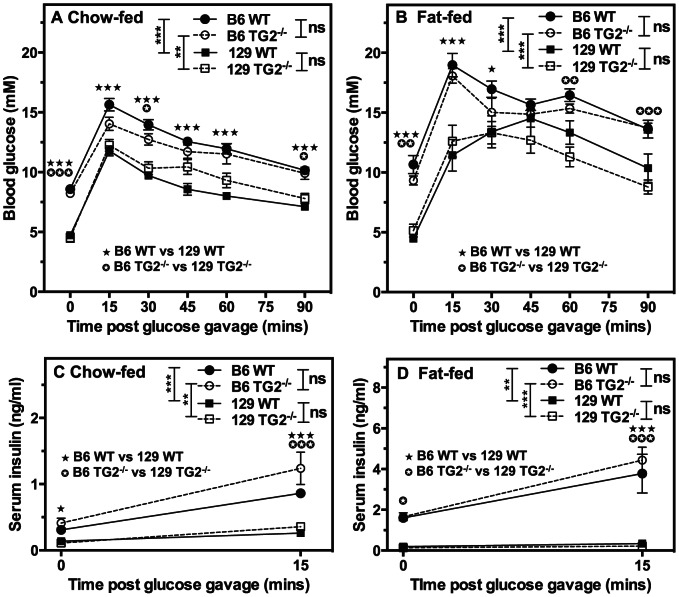
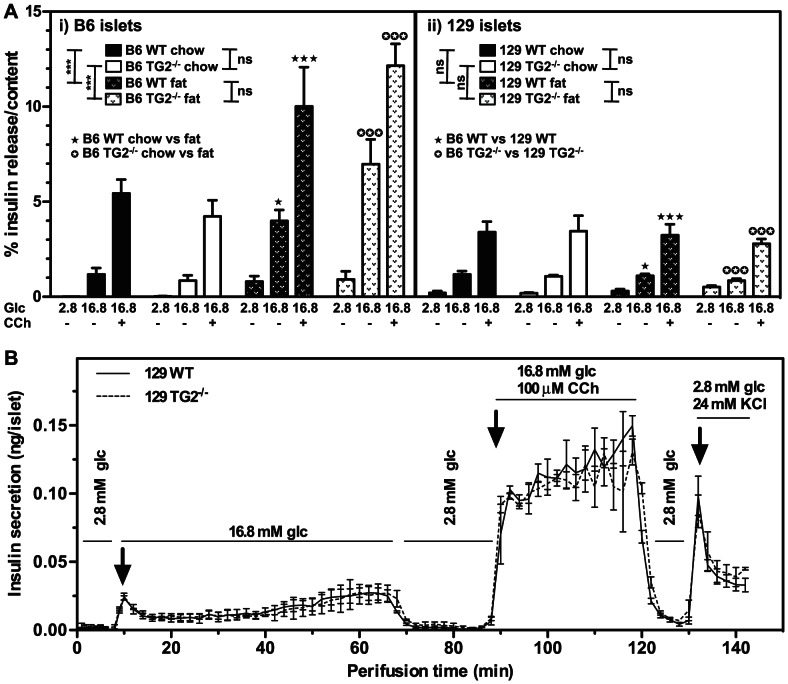
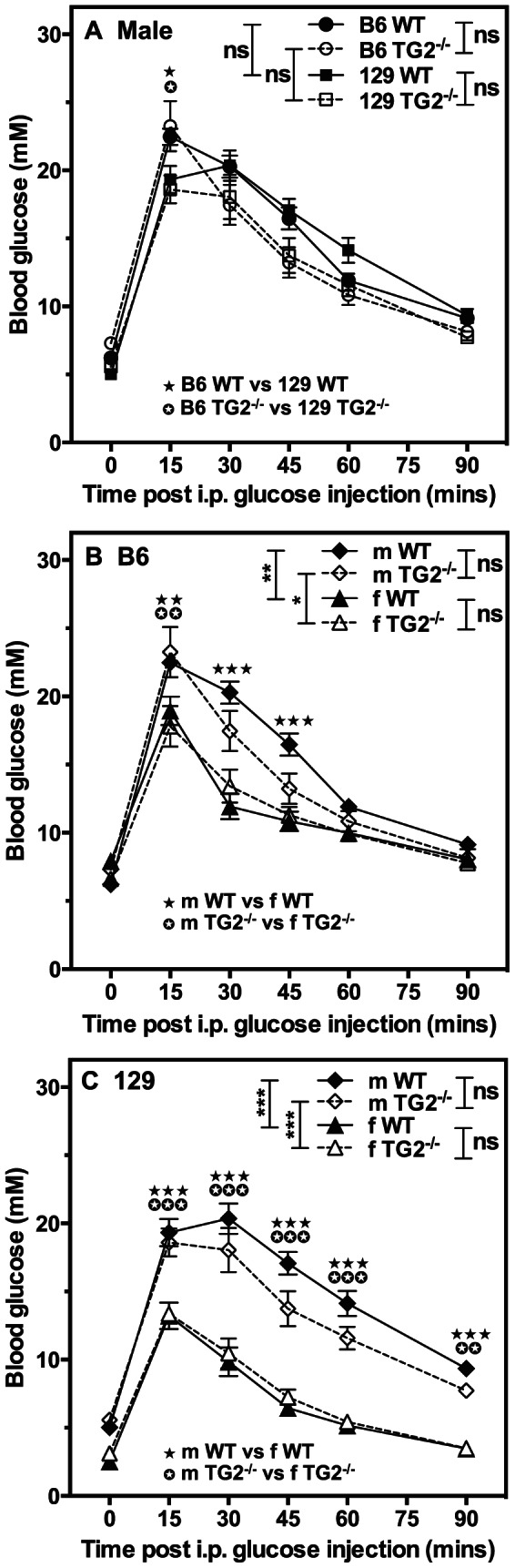
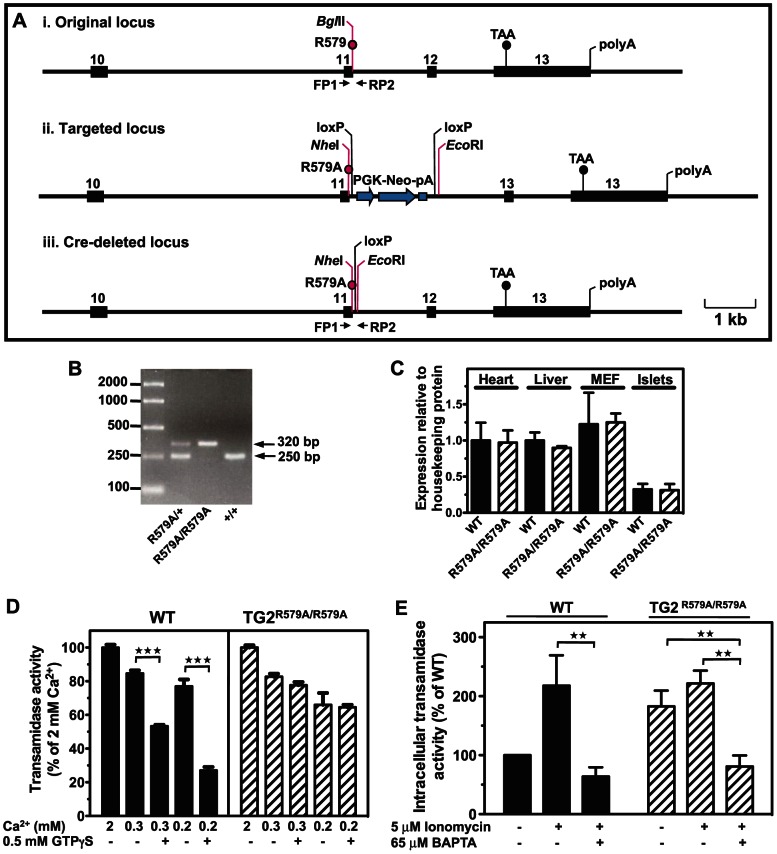
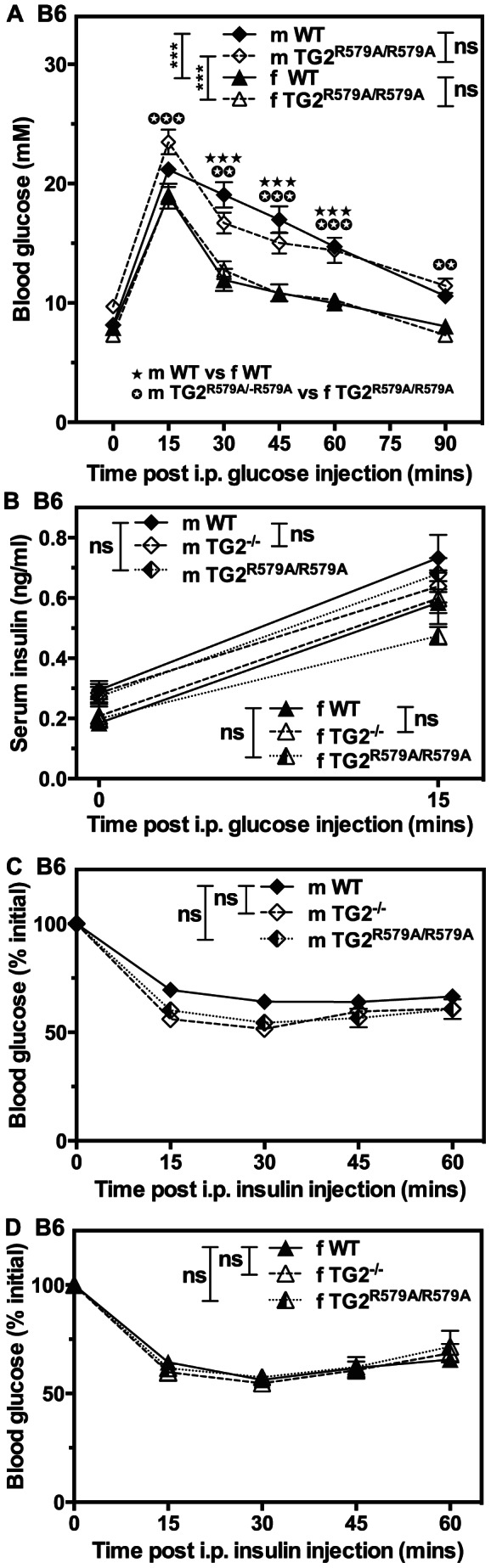
Transglutaminase type 2 (TG2) has been reported to be a candidate gene for maturity onset diabetes of the young (MODY) because three different mutations that impair TG2 transamidase activity have been found in 3 families with MODY. TG2 null (TG2(-/-)) mice have been reported to be glucose intolerant and have impaired glucose-stimulated insulin secretion (GSIS). Here we rigorously evaluated the role of TG2 in glucose metabolism using independently generated murine models of genetic TG2 disruption, which show no compensatory enhanced expression of other TGs in pancreatic islets or other tissues. First, we subjected chow- or fat-fed congenic SV129 or C57BL/6 wild type (WT) and TG2(-/-) littermates, to oral glucose gavage. Blood glucose and serum insulin levels were similar for both genotypes. Pancreatic islets isolated from these animals and analysed in vitro for GSIS and cholinergic potentiation of GSIS, showed no significant difference between genotypes. Results from intraperitoneal glucose tolerance tests (GTTs) and insulin tolerance tests (ITTs) were similar for both genotypes. Second, we directly investigated the role of TG2 transamidase activity in insulin secretion using a coisogenic model that expresses a mutant form of TG2 (TG2(R579A)), which is constitutively active for transamidase activity. Intraperitoneal GTTs and ITTs revealed no significant differences between WT and TG2(R579A/R579A) mice. Given that neither deletion nor constitutive activation of TG2 transamidase activity altered basal responses, or responses to a glucose or insulin challenge, our data indicate that glucose homeostasis in mice is TG2 independent, and question a link between TG2 and diabetes.
Conflict of interest statement
Figures
References
-
- Ledermann HM (1995) Maturity-onset diabetes of the young (MODY) at least ten times more commin in Europe than previously assumed? Diabetologia 38: 1482. - PubMed
-
- Shields BM, Hicks S, Shepherd MH, Colclough K, Hattersley AT, et al. (2010) Maturity-onset diabetes of the young (MODY): how many cases are we missing? Diabetologia 53: 2504–2508. - PubMed
-
- Bernassola F, Federici M, Corazzari M, Terrinoni A, Hribal ML, et al. (2002) Role of transglutaminase 2 in glucose tolerance: knockout mice studies and putative mutation in a MODY patient. FASEB J 16: 1371–1378. - PubMed
-
- Porzio O, Massa O, Cunsolo V, Colombo C, Malaponti M, et al. (2007) Missense mutations in the TGM2 gene encoding Transglutaminase 2 are found in patients with early-onset Type 2 diabetes. Hum Mutat 28: 1150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
