

Retroviral restriction factor APOBEC3G delays the initiation of DNA synthesis by HIV-1 reverse transcriptase
- PMID: 23717565
- PMCID: PMC3662766
- DOI: 10.1371/journal.pone.0064196
Retroviral restriction factor APOBEC3G delays the initiation of DNA synthesis by HIV-1 reverse transcriptase
Abstract
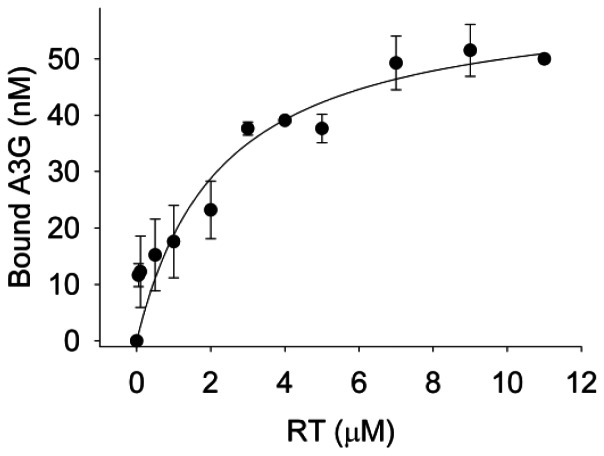
It is well established that the cytosine deaminase APOBEC3G can restrict HIV-1 virions in the absence of the virion infectivity factor (Vif) by inducing genome mutagenesis through deamination of cytosine to uracil in single-stranded HIV-1 (-)DNA. However, whether APOBEC3G is able to restrict HIV-1 using a deamination-independent mode remains an open question. In this report we use in vitro primer extension assays on primer/templates that model (-)DNA synthesis by reverse transcriptase from the primer binding site (PBS) and within the protease gene of HIV-1. We find that APOBEC3G is able to decrease the initiation of DNA synthesis by reverse transcriptase approximately 2-fold under conditions where reverse transcriptase is in excess to APOBEC3G, as found in HIV-1 virions. However, the delay in the initiation of DNA synthesis on RNA templates up to 120 nt did not decrease the total amount of primer extended after extended incubation unless the concentration of reverse transcriptase was equal to or less than that of APOBEC3G. By determining apparent Kd values of reverse transcriptase and APOBEC3G for the primer/templates and of reverse transcriptase binding to APOBEC3G we conclude that APOBEC3G is able to decrease the efficiency of reverse transcriptase-mediated DNA synthesis by binding to the RNA template, rather than by physically interacting with reverse transcriptase. All together the data support a model in which this deamination-independent mode of APOBEC3G would play a minor role in restricting HIV-1. We propose that the deamination-independent inhibition of reverse transcriptase we observed can be a mechanism used by APOBEC3G to slow down proviral DNA formation and increase the time in which single-stranded (-)DNA is available for deamination by APOBEC3G, rather than a direct mechanism used by APOBEC3G for HIV-1 restriction.
Conflict of interest statement
Figures





References
-
- Sheehy AM, Gaddis NC, Choi JD, Malim MH (2002) Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 418: 646–650. - PubMed
-
- Sheehy AM, Gaddis NC, Malim MH (2003) The antiretroviral enzyme APOBEC3G is degraded by the proteasome in response to HIV-1 Vif. Nat Med 9: 1404–1407. - PubMed
-
- Yu X, Yu Y, Liu B, Luo K, Kong W, et al. (2003) Induction of APOBEC3G ubiquitination and degradation by an HIV-1 Vif-Cul5-SCF complex. Science 302: 1056–1060. - PubMed
-
- Marin M, Rose KM, Kozak SL, Kabat D (2003) HIV-1 Vif protein binds the editing enzyme APOBEC3G and induces its degradation. Nat Med 9: 1398–1403. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
