

Control of angiogenesis by AIBP-mediated cholesterol efflux
- PMID: 23719382
- PMCID: PMC3760669
- DOI: 10.1038/nature12166
Control of angiogenesis by AIBP-mediated cholesterol efflux
Abstract
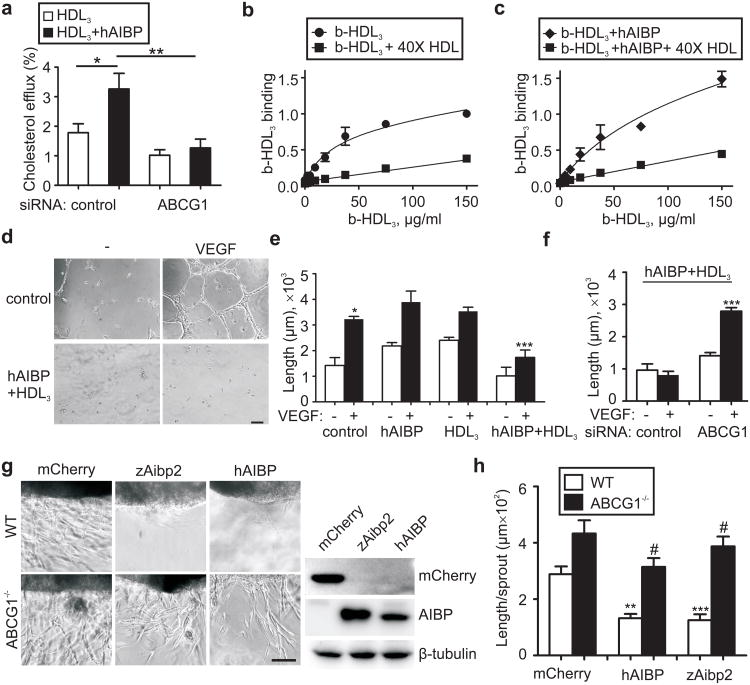
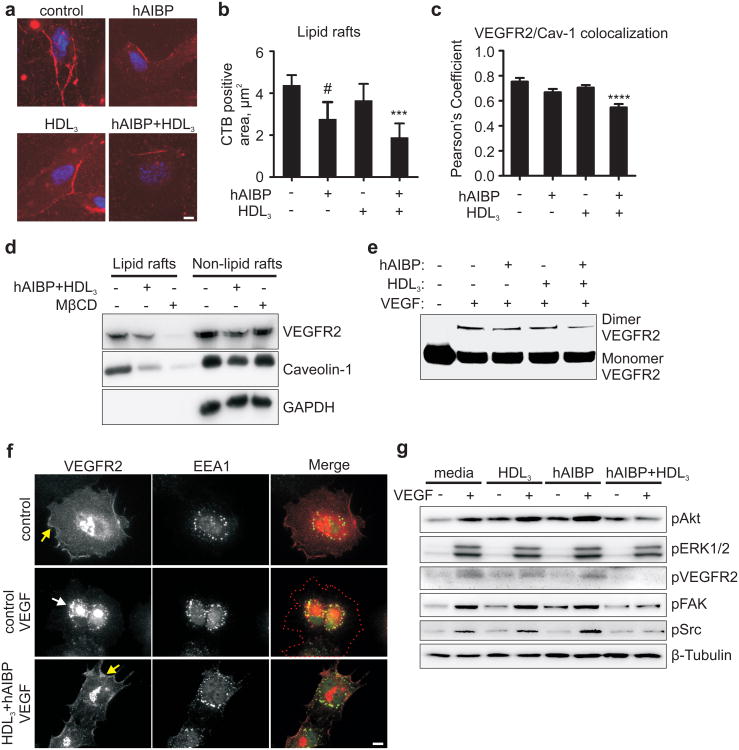
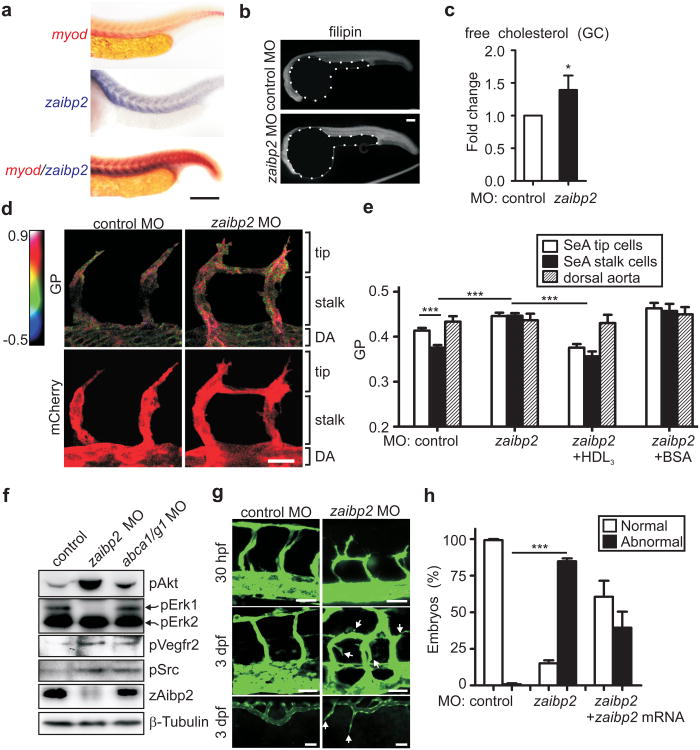
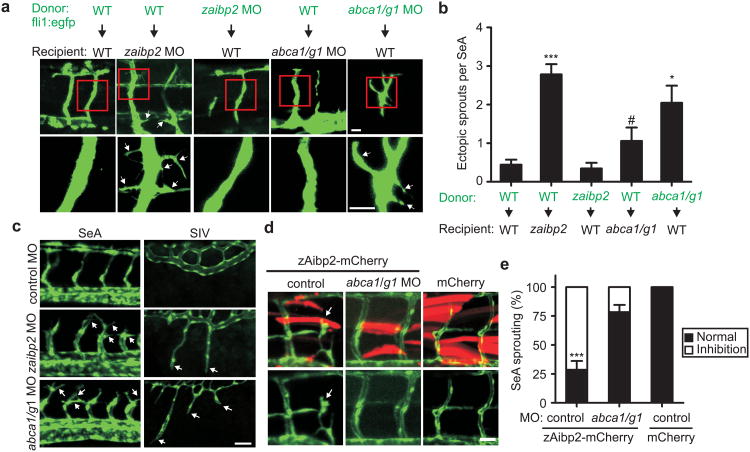
Cholesterol is a structural component of the cell and is indispensable for normal cellular function, although its excess often leads to abnormal proliferation, migration, inflammatory responses and/or cell death. To prevent cholesterol overload, ATP-binding cassette (ABC) transporters mediate cholesterol efflux from the cells to apolipoprotein A-I (apoA-I) and the apoA-I-containing high-density lipoprotein (HDL). Maintaining efficient cholesterol efflux is essential for normal cellular function. However, the role of cholesterol efflux in angiogenesis and the identity of its local regulators are poorly understood. Here we show that apoA-I binding protein (AIBP) accelerates cholesterol efflux from endothelial cells to HDL and thereby regulates angiogenesis. AIBP- and HDL-mediated cholesterol depletion reduces lipid rafts, interferes with VEGFR2 (also known as KDR) dimerization and signalling and inhibits vascular endothelial growth factor-induced angiogenesis in vitro and mouse aortic neovascularization ex vivo. Notably, Aibp, a zebrafish homologue of human AIBP, regulates the membrane lipid order in embryonic zebrafish vasculature and functions as a non-cell-autonomous regulator of angiogenesis. aibp knockdown results in dysregulated sprouting/branching angiogenesis, whereas forced Aibp expression inhibits angiogenesis. Dysregulated angiogenesis is phenocopied in Abca1 (also known as Abca1a) Abcg1-deficient embryos, and cholesterol levels are increased in Aibp-deficient and Abca1 Abcg1-deficient embryos. Our findings demonstrate that secreted AIBP positively regulates cholesterol efflux from endothelial cells and that effective cholesterol efflux is critical for proper angiogenesis.
Conflict of interest statement
The authors declare no conflicts of interests.
Figures
Comment in
-
Targeted cholesterol efflux.Cell Cycle. 2013 Nov 1;12(21):3345-6. doi: 10.4161/cc.26401. Epub 2013 Sep 13. Cell Cycle. 2013. PMID: 24036659 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
