

Conflicts between local and global spatial frameworks dissociate neural representations of the lateral and medial entorhinal cortex
- PMID: 23719794
- PMCID: PMC3747988
- DOI: 10.1523/JNEUROSCI.0946-13.2013
Conflicts between local and global spatial frameworks dissociate neural representations of the lateral and medial entorhinal cortex
Erratum in
- J Neurosci. 2013 Aug 7;33(32):13249
Abstract
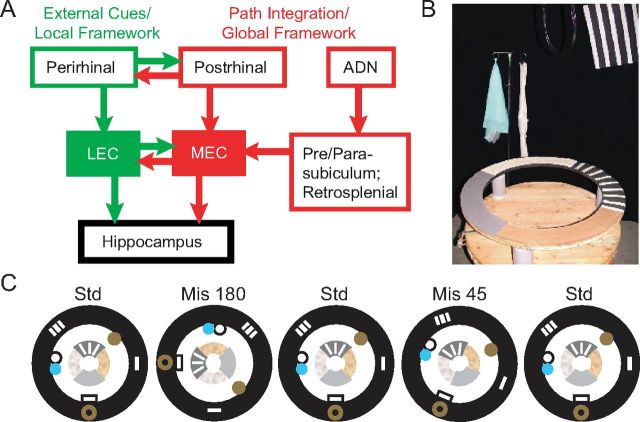
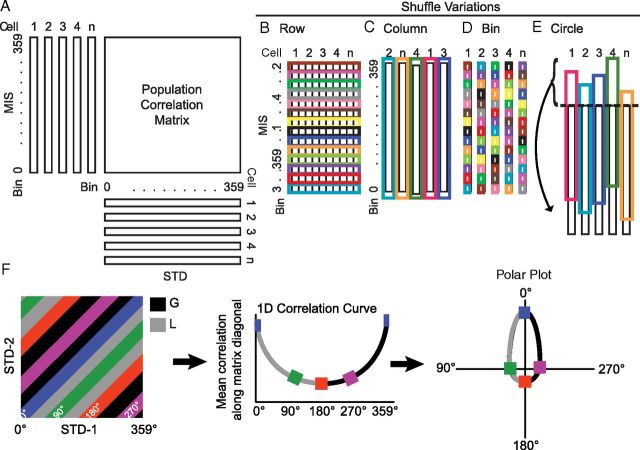
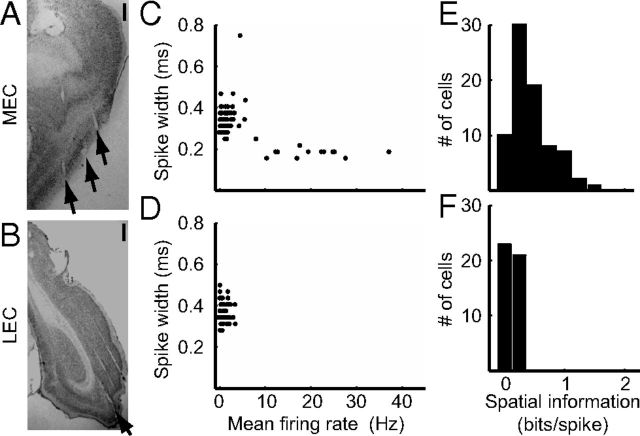
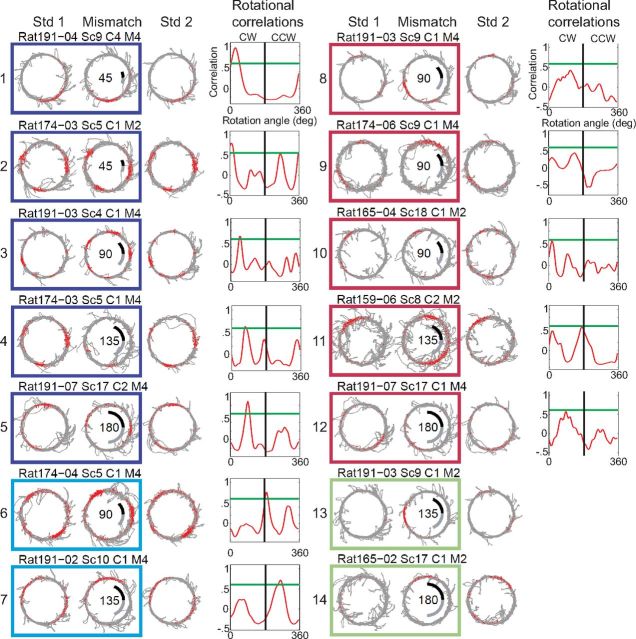
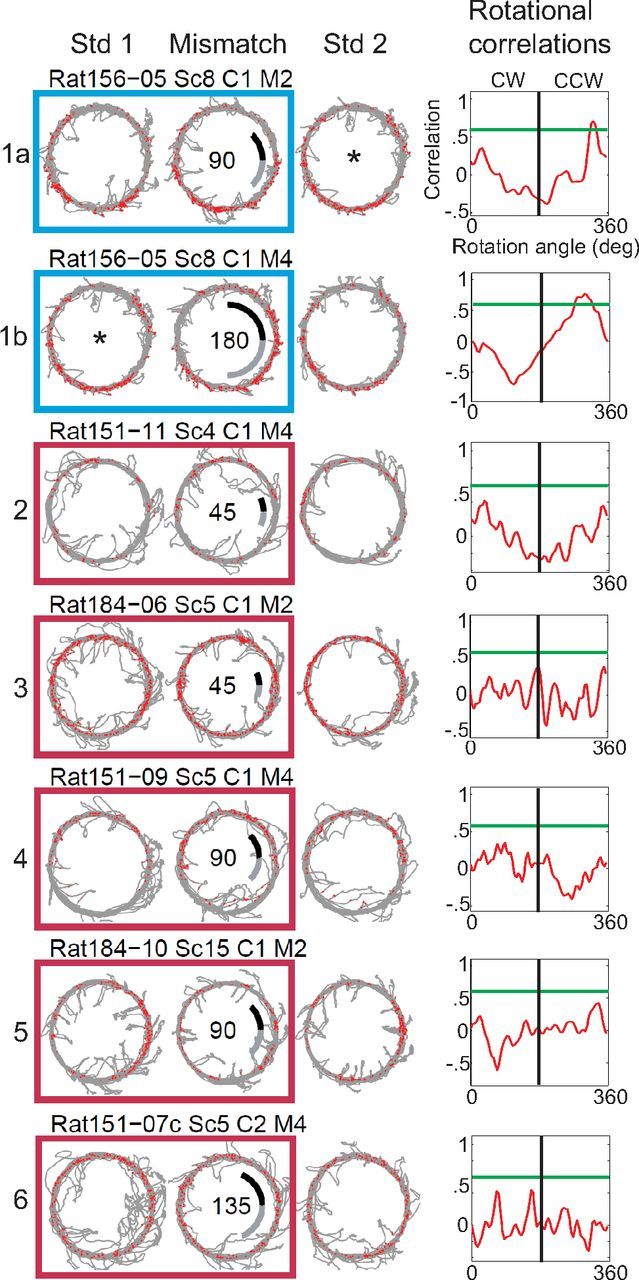
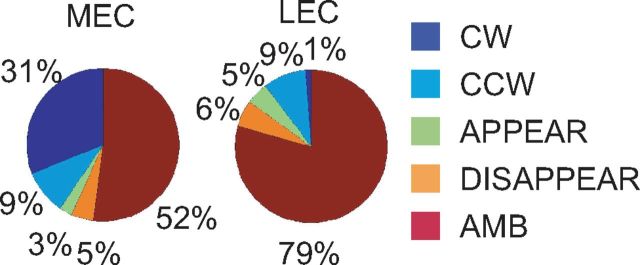
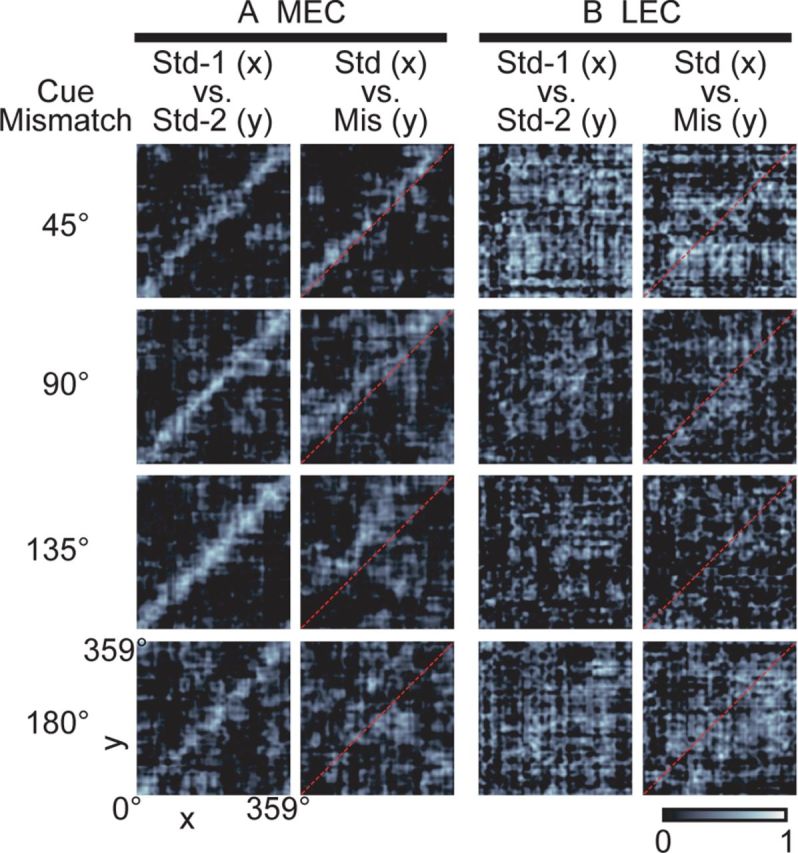
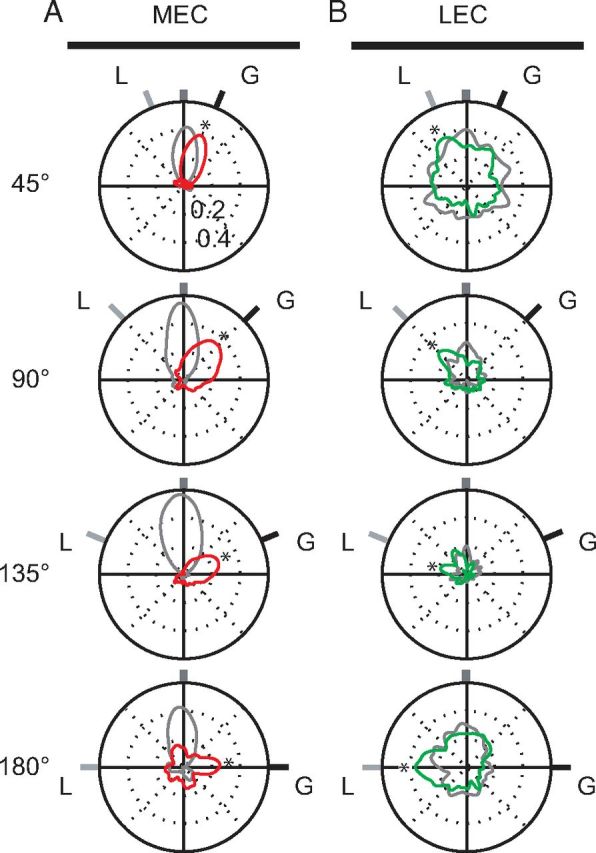
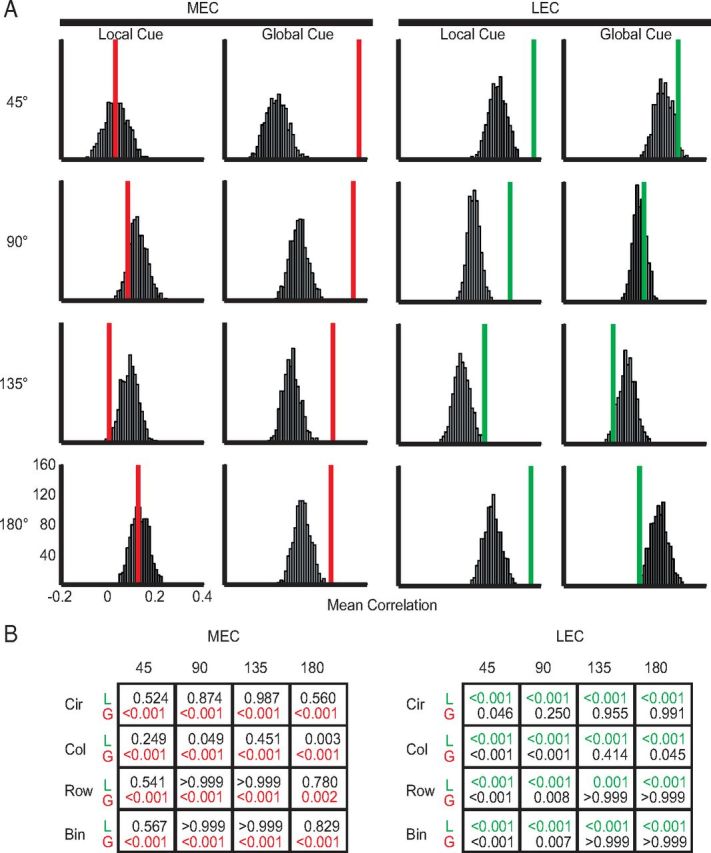
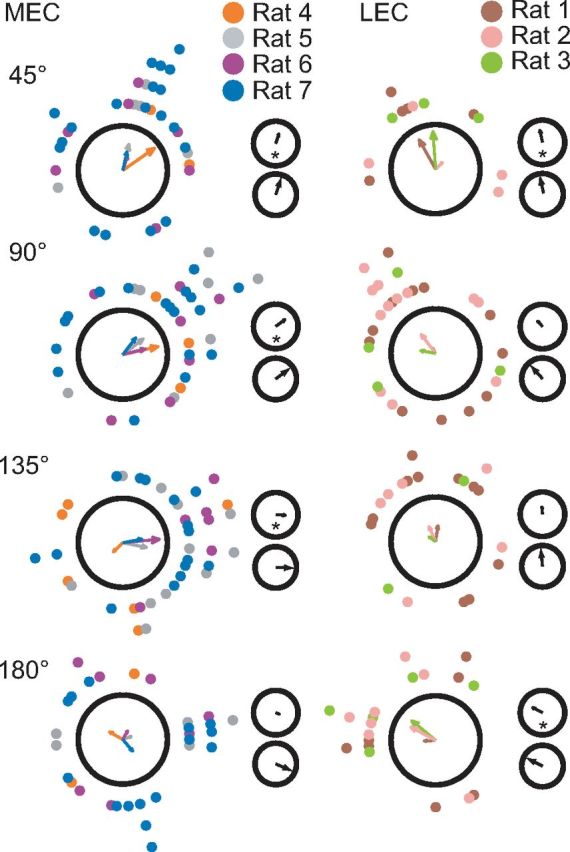
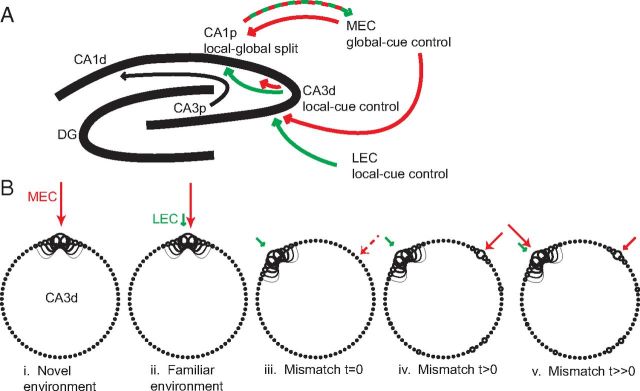
Manipulation of spatial reference frames is a common experimental tool to investigate the nature of hippocampal information coding and to investigate high-order processes, such as cognitive coordination. However, it is unknown how the hippocampus afferents represent the local and global reference frames of an environment. To address these issues, single units were recorded in freely moving rats with multi-tetrode arrays targeting the superficial layers of the lateral entorhinal cortex (LEC) and medial entorhinal cortex (MEC), the two primary cortical inputs to the hippocampus. Rats ran clockwise laps around a circular track partitioned into quadrants covered by different textures (the local reference frame). The track was centered in a circular environment with distinct landmarks on the walls (the global reference frame). Here we demonstrate a novel dissociation between MEC and LEC in that the global frame controlled the MEC representation and the local frame controlled the LEC representation when the reference frames were rotated in equal, but opposite, directions. Consideration of the functional anatomy of the hippocampal circuit and popular models of attractor dynamics in CA3 suggests a mechanistic explanation of previous data showing a dissociation between the CA3 and CA1 regions in their responses to this local-global conflict. Furthermore, these results are consistent with a model of the LEC providing the hippocampus with the external sensory content of an experience and the MEC providing the spatial context, which combine to form conjunctive codes in the hippocampus that form the basis of episodic memory.
Figures
Comment in
-
Location memory: separate cortical coding for distal and local cues.Curr Biol. 2013 Aug 19;23(16):R685-7. doi: 10.1016/j.cub.2013.07.018. Curr Biol. 2013. PMID: 23968923
References
-
- Brown JE, Skaggs WE. Concordant and discordant coding of spatial location in populations of hippocampal CA1 pyramidal cells. J Neurophysiol. 2002;88:1605–1613. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous