

Molecular mechanism of TRP channels
- PMID: 23720286
- PMCID: PMC3775668
- DOI: 10.1002/cphy.c120001
Molecular mechanism of TRP channels
Abstract
Transient receptor potential (TRP) channels are cellular sensors for a wide spectrum of physical and chemical stimuli. They are involved in the formation of sight, hearing, touch, smell, taste, temperature, and pain sensation. TRP channels also play fundamental roles in cell signaling and allow the host cell to respond to benign or harmful environmental changes. As TRP channel activation is controlled by very diverse processes and, in many cases, exhibits complex polymodal properties, understanding how each TRP channel responds to its unique forms of activation energy is both crucial and challenging. The past two decades witnessed significant advances in understanding the molecular mechanisms that underlie TRP channels activation. This review focuses on our current understanding of the molecular determinants for TRP channel activation.
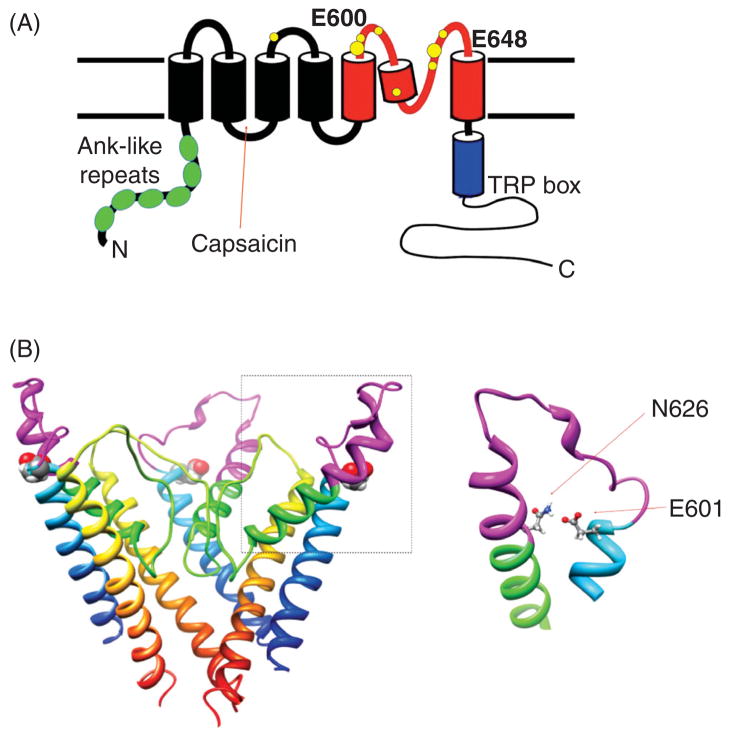
Figures
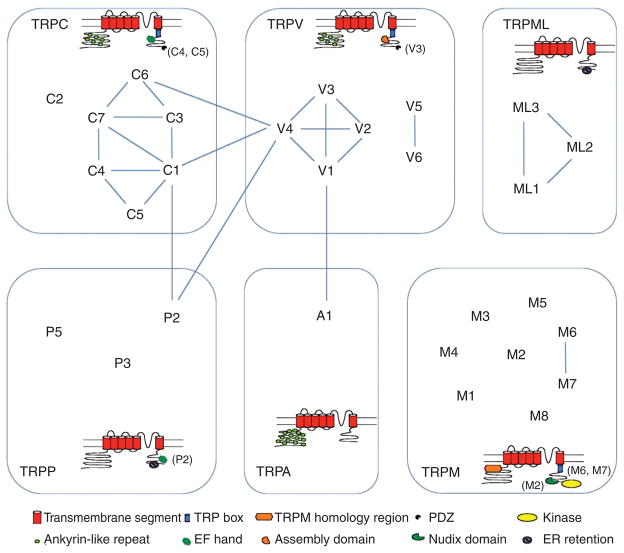
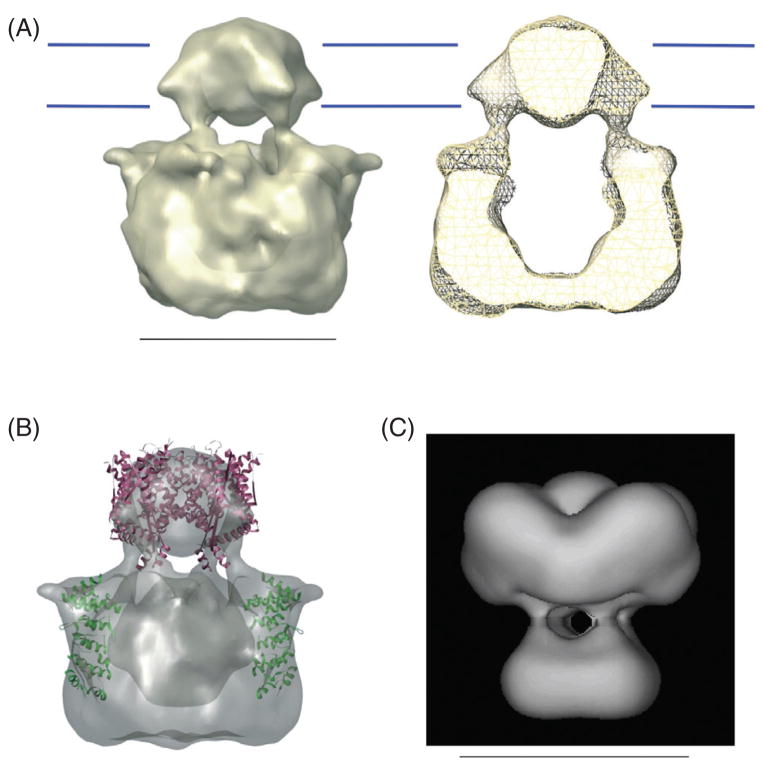
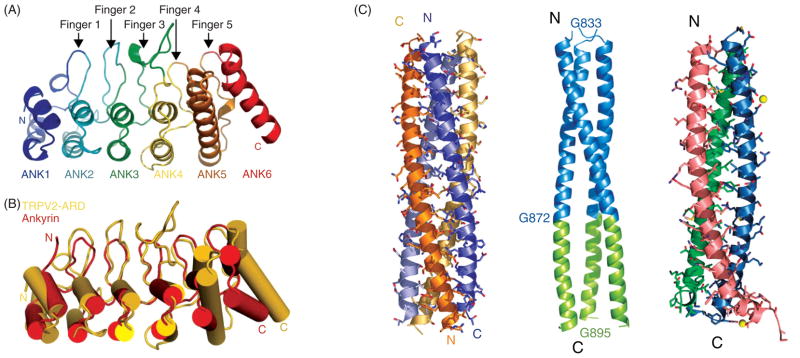
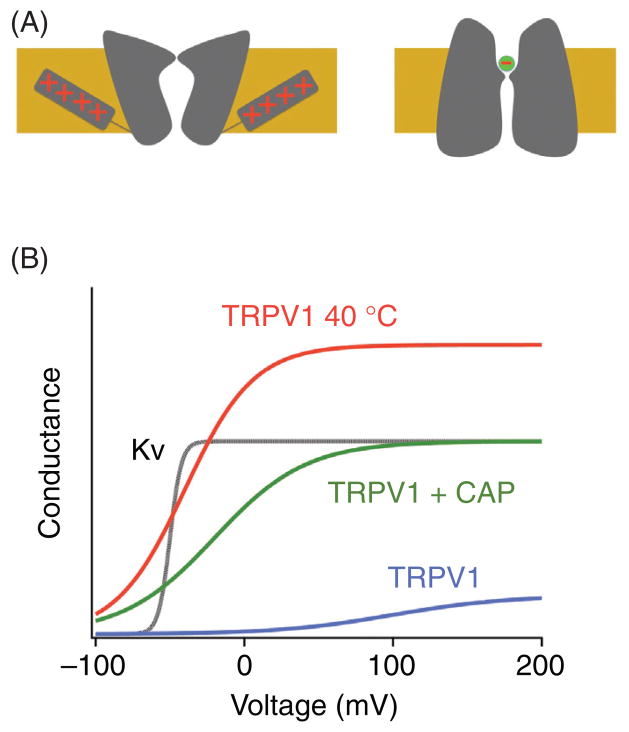
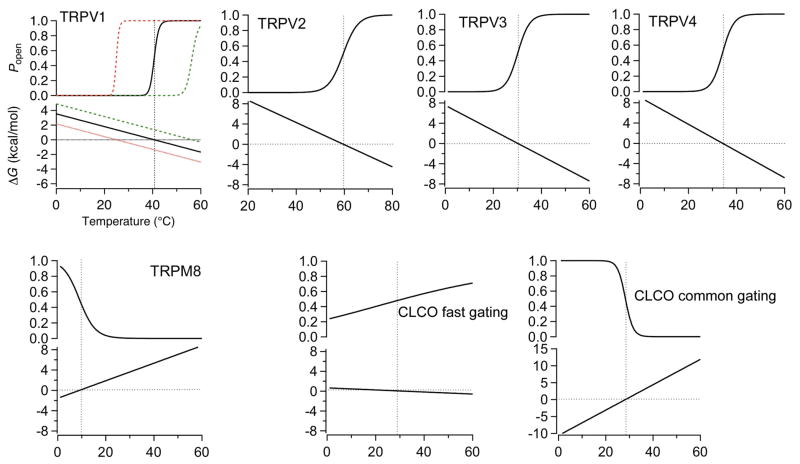
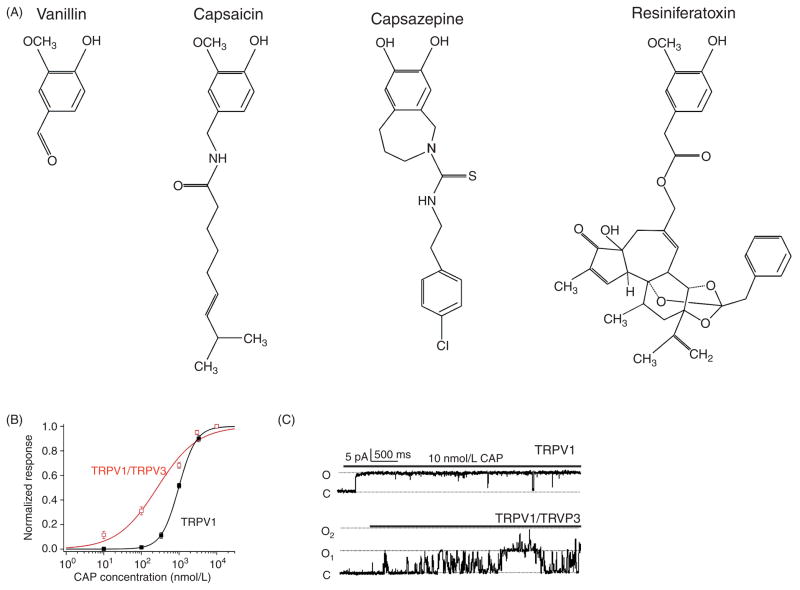
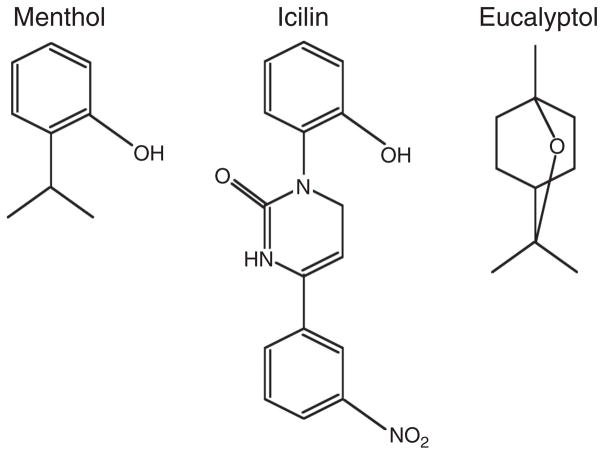
References
-
- Arniges M, Fernandez-Fernandez JM, Albrecht N, Schaefer M, Valverde MA. Human TRPV4 channel splice variants revealed a key role of ankyrin domains in multimerization and trafficking. J Biol Chem. 2006;281:1580–1586. - PubMed
-
- Bandell M, Dubin AE, Petrus MJ, Orth A, Mathur J, Hwang SW, Patapoutian A. High-throughput random mutagenesis screen reveals TRPM8 residues specifically required for activation by menthol. Nat Neurosci. 2006;9:493–500. - PubMed
Further Reading
-
- Hille Bertil. Ion Channels of Excitable Membranes. 3. Sunderland: Sinauer Associates, Inc; 2001.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
