

Review
doi: 10.1002/cphy.c120019.
Physiology of cholangiocytes
Affiliations
- PMID: 23720296
- PMCID: PMC3831353
- DOI: 10.1002/cphy.c120019
Item in Clipboard
Review
Physiology of cholangiocytes
Compr Physiol.
2013 Jan.
Abstract
Cholangiocytes are epithelial cells that line the intra- and extrahepatic ducts of the biliary tree. The main physiologic function of cholangiocytes is modification of hepatocyte-derived bile, an intricate process regulated by hormones, peptides, nucleotides, neurotransmitters, and other molecules through intracellular signaling pathways and cascades. The mechanisms and regulation of bile modification are reviewed herein.
Figures
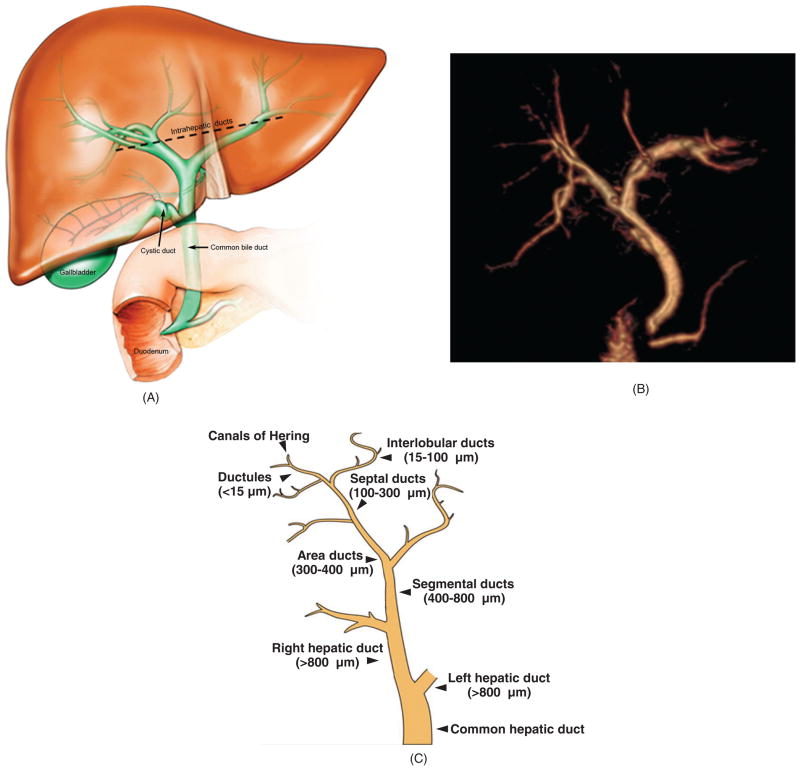
Biliary tree architecture. (A) The biliary tree is a three-dimensional network of interconnecting tubular conduits consisting of an intrahepatic (above dashed black line) and an extrahepatic (below dashed black line) portion. (B) Magnetic resonance cholangiogram of human biliary tree (three-dimensional reconstruction). Also seen at the lower portion of the extrahepatic biliary tree are the pancreatic duct and the duodenum, the latter being where biliary and pancreatic contents drain. The gallbladder is surgically absent. (C) The human biliary tree comprises conduits of different sizes extending from the canals of Hering, through which primary or canalicular bile enters the biliary tree, to the terminus of the common bile duct (i.e., where it empties into the duodenum). [B: courtesy of Dr. Naoki Takahashi and Dr. Joel Glockner, Mayo Clinic; C: Reproduced from Masyuk et al. (183), with permission].
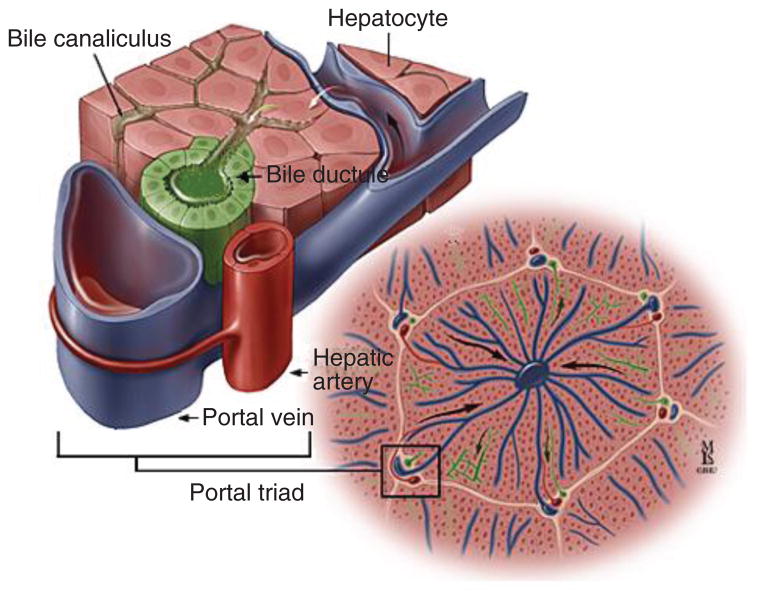
Portal triad. Consists of a biliary (represented here as “bile ductule”), hepatic arterial, and portal venous component. [Courtesy of Dr. Anthony Kalloo, Johns Hopkins Gastroenterology & Hepatology (www.hopkins-gi.org )].

Transmission electron micrographs of rat intrahepatic bile duct cross-sections. (A) Large bile duct lined by 11 cholangiocytes, each with an apical and basolateral plasma membrane and demarcated by tight junctions (TJ) located near the APM. Microvilli on the apical plasma membrane significantly increase the cholangiocyte surface area. (B) Higher power magnification of large bile duct showing partial long view and axial section (black arrows) of primary cilia. [A: reproduced from Masyuk et al. (183), with permission].
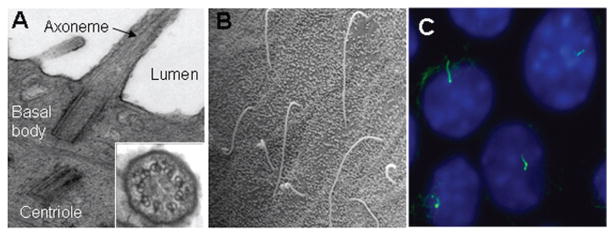
Cholangiocyte primary cilia. (A) Transmission electron microscopy micrograph of primary cilia extending from the cholangiocyte apical plasma membrane and into the ductal lumen with insert showing the 9+0 pattern of the ciliary axoneme. (B) Scanning electron microscopy images of primary cilia in large rat bile duct. (C) Immunofluorescence confocal microscopy image of primary cilia in normal mouse cholangiocyte cell line 10 to 14 days after confluence. In C, cilia were stained with acetylated α-tubulin (green) and nuclei were stained with DAPI (blue). [A and C: reproduced from Masyuk et al. (189), with permission].
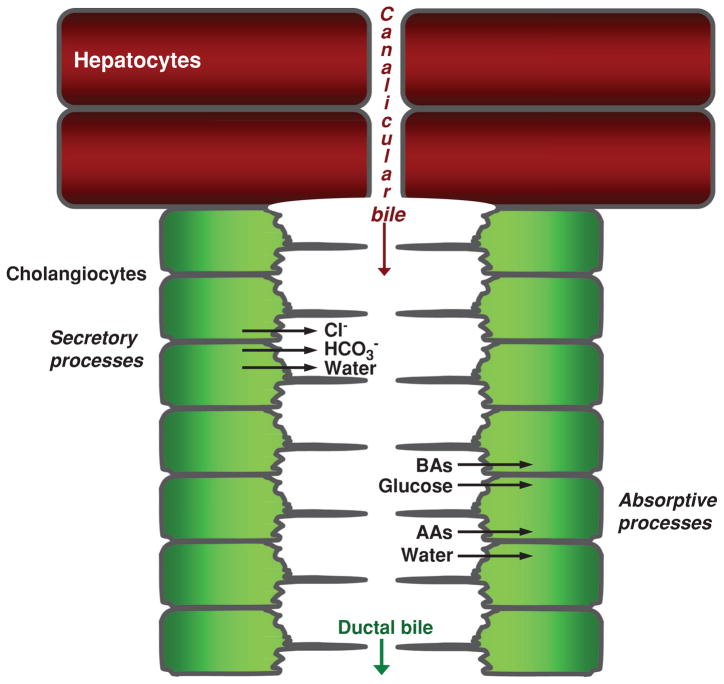
Overview of bile formation and modification. Hepatocytes initiate bile formation by secreting primary bile, composed primarily of water, solutes, and ions, into canaliculi. As canalicular bile flows along the biliary tree, it is subjected to cholangiocyte secretory and absorptive processes, thus resulting in modified ductal bile. Secretion of Cl−, HCO3−, and water and absorption of bile acids (BAs), glucose, amino acids (AAs), and water are the major transport processes determining the chemical composition of ductal bile. [Adapted from Masyuk et al. (183), with permission].
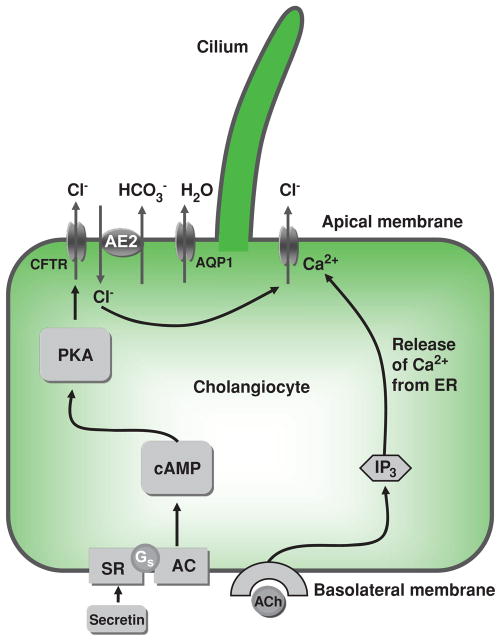
Mechanisms of ductal bile modification. According to a widely accepted model, secretin induces HCO3−-rich fluid secretion from cholangiocytes via the cyclic adenosine 3′,5′-monophosphate-protein kinase A (cAMP-PKA) signaling pathway by activation of the apical cystic fibrosis transmembrane conductance regulator (CFTR) Cl−channel resulting in extrusion of Cl− ions, which in turn stimulate the Cl−/HCO3− exchanger AE2 and subsequent secretion of HCO3−. Secreted HCO3− ions drive passive aquaporin 1 (AQP1)-mediated movement of water in response to established osmotic gradients. An alternative mechanism of Cl– efflux is also shown, consisting of acetylcholine (ACh) binding to basolateral M3 muscarinic receptors, which leads to increases in inositol trisphosphate (IP3), which binds to (types I and II) IP3Rs, causing release of Ca2+ and activation of apical Cl− secretion. AC, adenylate cyclase; Gs, Gs protein; PKA, protein kinase A; SR, secretin receptor. [Adapted from Masyuk et al. (183), with permission].
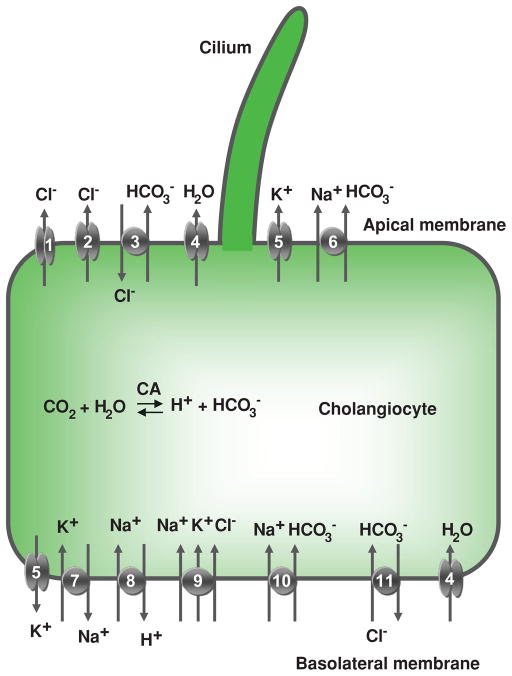
Cholangiocyte secretion. Located at the cholangiocyte apical plasma membrane are: (1) cyclic adenosine 3′,5′-monophosphate (cAMP)-regulated Cl− channel (cystic fibrosis transmembrane conductance regulator), (2) Ca2+-regulated Cl− channel, (3) Na+-independent Cl−/HCO3− exchanger, (4) water channel, (5) K+ channel (SK2), and (6) Na+-HCO3− cotransporter (in mouse). At the basolateral plasma membrane, cholangiocytes express (5) K+ channels (SK2 and IK-1), (7) Na+/K+-ATPase, (8) Na+/H+ exchanger, (9) Na+- K+-Cl− cotransporter, (10) Na+-HCO3− cotransporter (in rat), (11) Na+-dependent Cl−/HCO3− exchanger (in humans), and (4) water channels (AQP1 and AQP4). Details regarding each of these components of cholangiocyte secretion are found in the text. CA, carbonic anhydrase. [Adapted from Masyuk et al. (183), with permission].
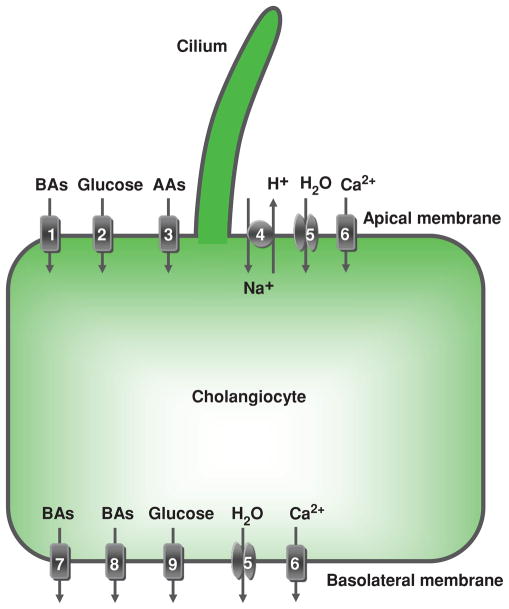
Cholangiocyte absorption. Located at the cholangiocyte apical plasma membrane are: (1) Na+-dependent bile acid transporter (ASBT), (2) Na+-dependent glucose transporter, (3) unidentified Na+-dependent and Na+-independent amino acid transporters, (4) Na+/H+ exchanger, (5) a water channel (AQP1), and (6) plasma membrane Ca2+ pump (PMCA) (Na+/Ca2+ exchanger not shown due to inadequate data in cholangiocytes). At the basolateral plasma membrane, cholangiocytes express: bile acid transporters (7) t-ASBT and (8) Ostα-Ostβ, (9) glucose transporter, (5) water channels (AQP1 and AQP4), and (6) PMCA (speculated to be present on both the apical and basolateral membrane). These, as well as the secretory transport proteins, function in a coordinated manner to contribute to ductal bile modification by absorbing solutes and water from bile and transporting them out of the cholangiocyte. Details regarding each of these components of cholangiocyte absorption are found in the text. AAs, amino acids; BAs, bile acids. [Adapted from Masyuk et al. (183), with permission].
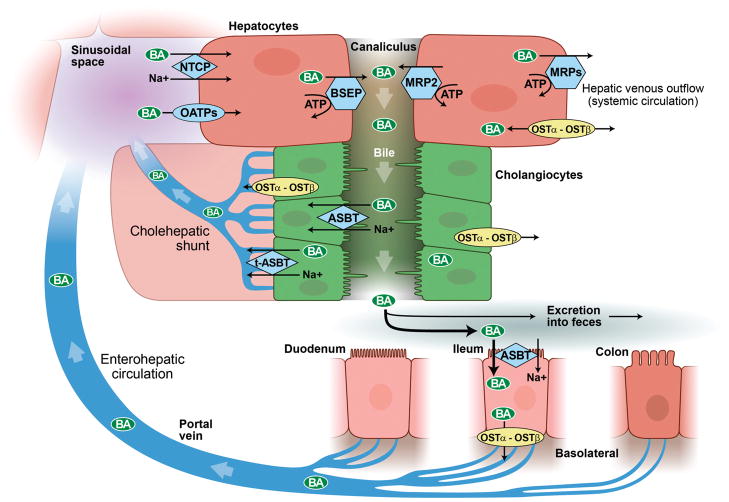
Enterohepatic circulation and cholehepatic shunt. Bile acids are synthesized by hepatocytes and subsequently secreted into canalicular bile by means of specialized hepatocyte canalicular membrane transporters. Canalicular bile drains into the biliary tree and is modified by the epithelial cells lining it, that is, cholangiocytes. Bile then drains into the proximal small bowel, that is, duodenum, and is metabolized by enteric bacteria. Approximately 95% of bile acids are reabsorbed in the terminal ileum and enter the portal vein to be recycled back to the liver via the enterohepatic circulation. Once in the sinusoids of the liver, bile acids can be taken up by hepatocytes and secreted back into bile. A fraction of (unconjugated) bile acids in the biliary tree are taken up by cholangiocytes at the apical membrane (i.e., prior to reaching the small intestine) and returned to the liver sinusoids via the cholehepatic shunt. (With permission of Mayo Foundation for Medical Education and Research. All Rights Reserved.)
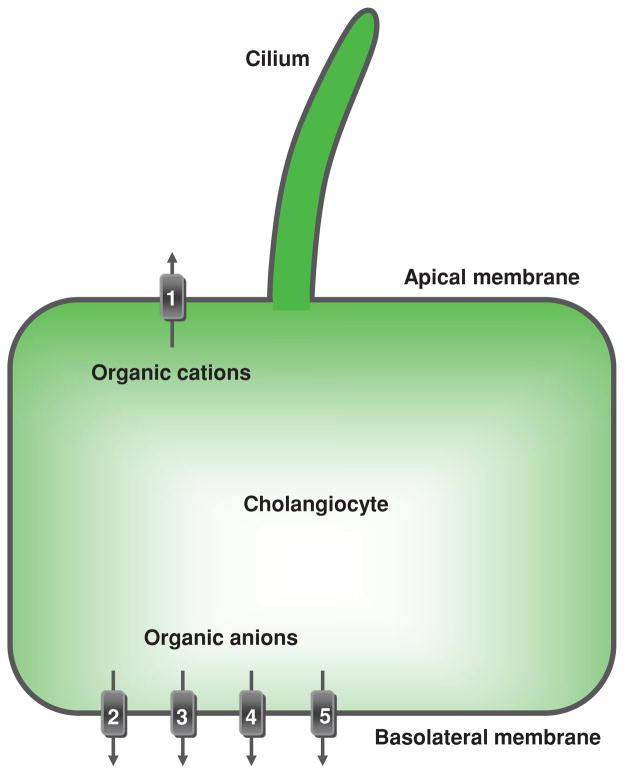
Cholangiocyte transport of organic anions and cations. Located at the cholangiocyte apical plasma membrane is (1) multidrug resistance 1 protein that excretes organic cations into bile. Located at the basolateral plasma membrane are transporters effectuating efflux of organic anions from cholangiocytes into the peribiliary vascular plexus: multidrug-associated proteins (2) MPR3/Mpr3 and (3) MPR4, (4) organic anion transporting polypeptide 3, and (5) organic solute transporter Ostα-Ostβ. TJ, tight junction. [Adapted from Masyuk et al. (183), with permission].
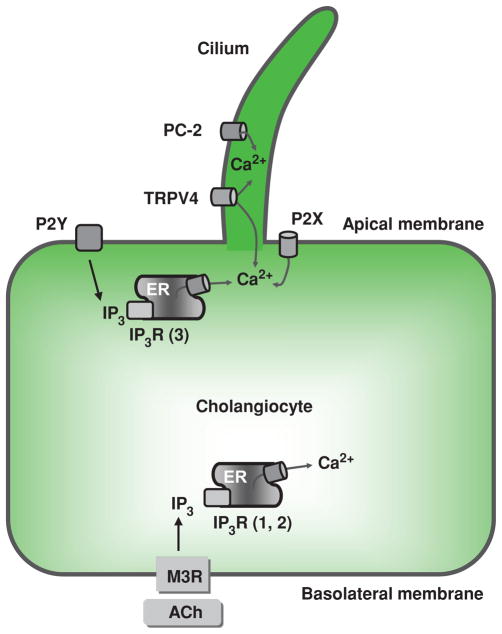
Ca2+ signaling in cholangiocytes. At the cholangiocyte apical plasma membrane, Ca2+ signaling is initiated by activation of the G-protein-coupled receptors (e.g., P2Y), resulting in production of inositol 1,4,5-trisphosphate (IP3). IP3, via the IP3 receptor (IP3R)-isoform 3, induces Ca2+ release from the endoplasmic reticulum (ER), increasing the intracellular Ca2+ concentration. Intracellular Ca2+ concentration is also increased as a result of entry of extracellular Ca2+ via apically and ciliary located PC-2, P2X, and TRPV4 that function as Ca2+ channels. Activation of basolaterally located muscarinic receptors (M3R) by acetylcholine (ACh) also results in production of IP3 that in turn releases Ca2+ from ER via the IP3R isoforms 1 and 2. TJ, tight junction. [Adapted from Masyuk et al. (183), with permission].
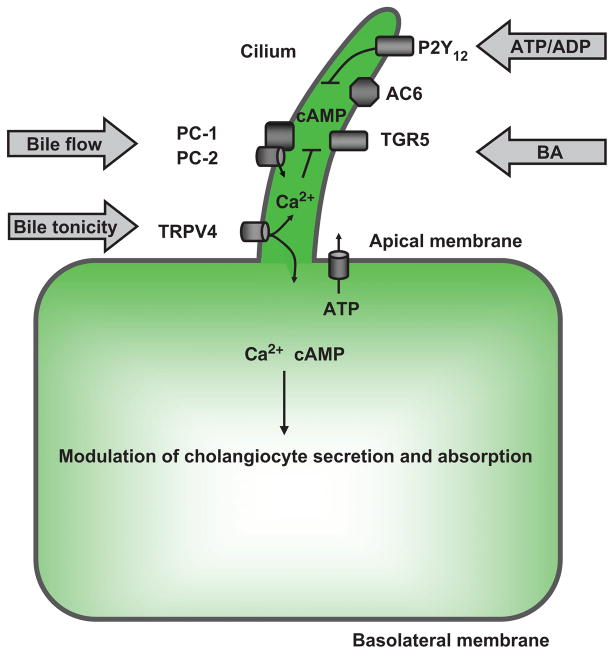
Cholangiocyte cilia in bile modification. A working model shows the potential involvement of mechano-, osmo-, and chemosensory functions of cholangiocyte cilia in intracellular signaling associated with ductal bile modification. In response to mechanical stimuli (e.g., bile flow), cilia-associated PC-1 and PC-2 form a functional complex allowing entry of extracellular Ca2+, which in turn suppresses cyclic adenosine 3′,5′-monophosphate (cAMP) concentration in cholangio-cytes via cilia-associated AC6. In response to osmostimuli (i.e., changes in bile tonicity), cilia-associated TRPV4 is activated when bile tonicity decreases or inhibited when bile tonicity increases, causing changes in intracellular Ca2+ concentration, which in turn affect cholangiocyte ATP release via unknown mechanisms. As an example of chemostimuli, biliary ATP, released by hepatocytes and/or cholangiocytes, as well as ATP degradation product ADP, inhibit ciliary cAMP signaling via cilia-associated P2Y12, whereas bile acids induce an increase in cAMP levels via cilia-associated bile acid receptor, TGR5. Intracellular cAMP and Ca2+ signaling induced by extracellular stimuli via cilia-associated mechanisms may affect apically and basolaterally located transport mechanisms depicted in Figures 7 and 9 resulting in increased or decreased cholangiocyte secretion or absorption. [Adapted from Masyuk et al. (183), with permission].
References
-
- Abbracchio MP, Burnstock G, Boeynaems JM, Barnard EA, Boyer JL, Kennedy C, Knight GE, Fumagalli M, Gachet C, Jacobson KA, Weisman GA. International Union of Pharmacology LVIII: Update on the P2Y G protein-coupled nucleotide receptors: from molecular mechanisms and pathophysiology to therapy. Pharmacol Rev. 2006;58:281–341. - PMC - PubMed
-
- Alper SL. Molecular physiology of SLC4 anion exchangers. Exp Physiol. 2006;91:153–161. - PubMed
-
- Alpini G, Baiocchi L, Glaser S, Ueno Y, Marzioni M, Francis H, Phinizy JL, Angelico M, Lesage G. Ursodeoxycholate and tauroursodeoxycholate inhibit cholangiocyte growth and secretion of BDL rats through activation of PKC alpha. Hepatology. 2002;35:1041–1052. - PubMed
-
- Alpini G, Franchitto A, Demorrow S, Onori P, Gaudio E, Wise C, Francis H, Venter J, Kopriva S, Mancinelli R, Carpino G, Stagnitti F, Ueno Y, Han Y, Meng F, Glaser S. Activation of alpha(1) -adrenergic receptors stimulate the growth of small mouse cholangiocytes via calcium-dependent activation of nuclear factor of activated T cells 2 and specificity protein 1. Hepatology. 2011;53:628–639. - PMC - PubMed
-
- Alpini G, Glaser S, Alvaro D, Ueno Y, Marzioni M, Francis H, Baiocchi L, Stati T, Barbaro B, Phinizy JL, Mauldin J, Lesage G. Bile acid depletion and repletion regulate cholangiocyte growth and secretion by a phosphatidylinositol 3-kinase-dependent pathway in rats. Gastroenterology. 2002;123:1226–1237. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
