

Aire-expressing thymic medullary epithelial cells originate from β5t-expressing progenitor cells
- PMID: 23720310
- PMCID: PMC3683726
- DOI: 10.1073/pnas.1301799110
Aire-expressing thymic medullary epithelial cells originate from β5t-expressing progenitor cells
Abstract
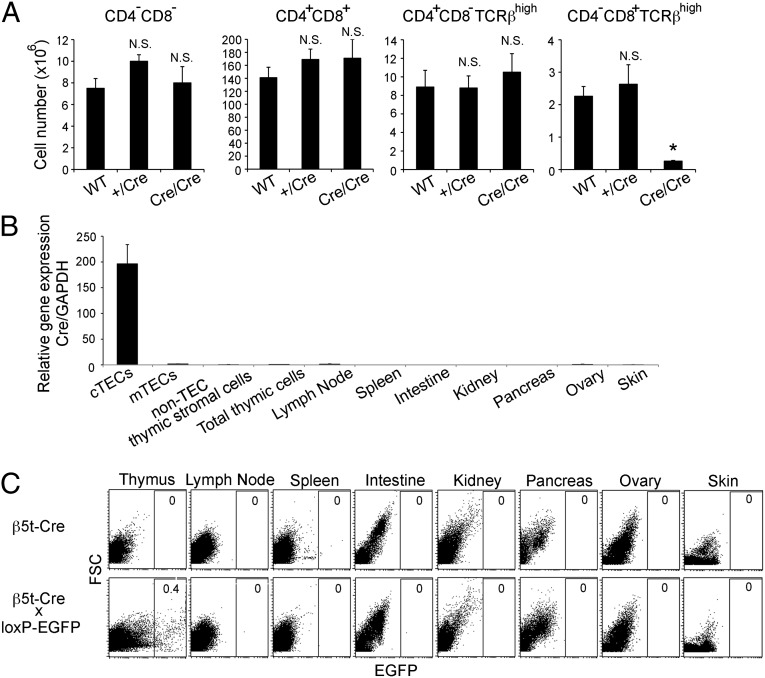
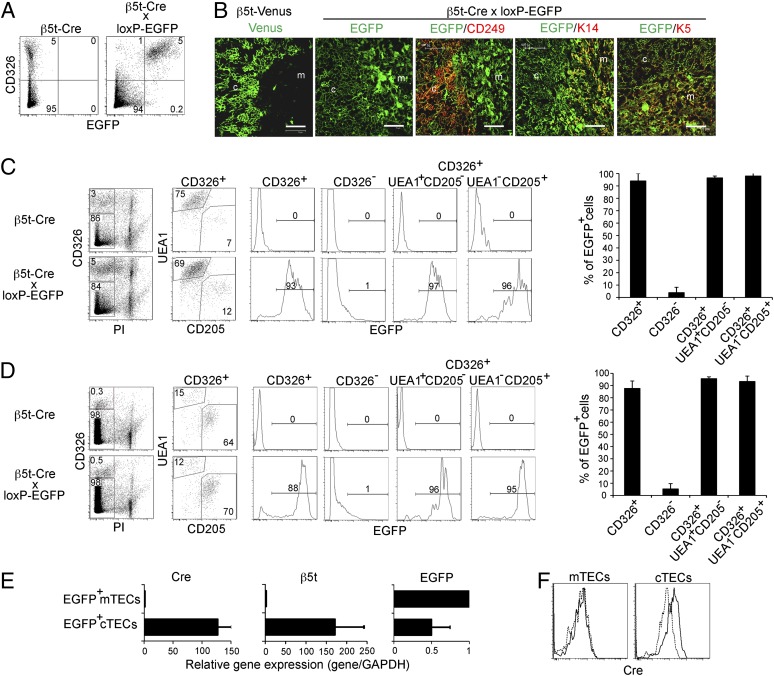
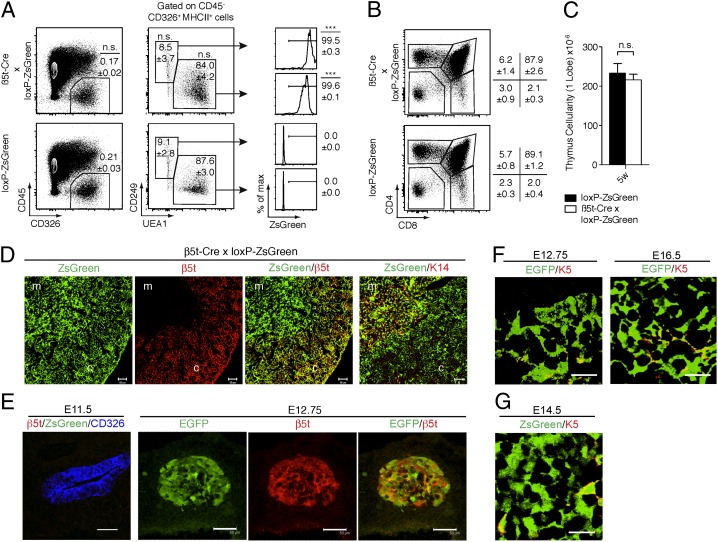
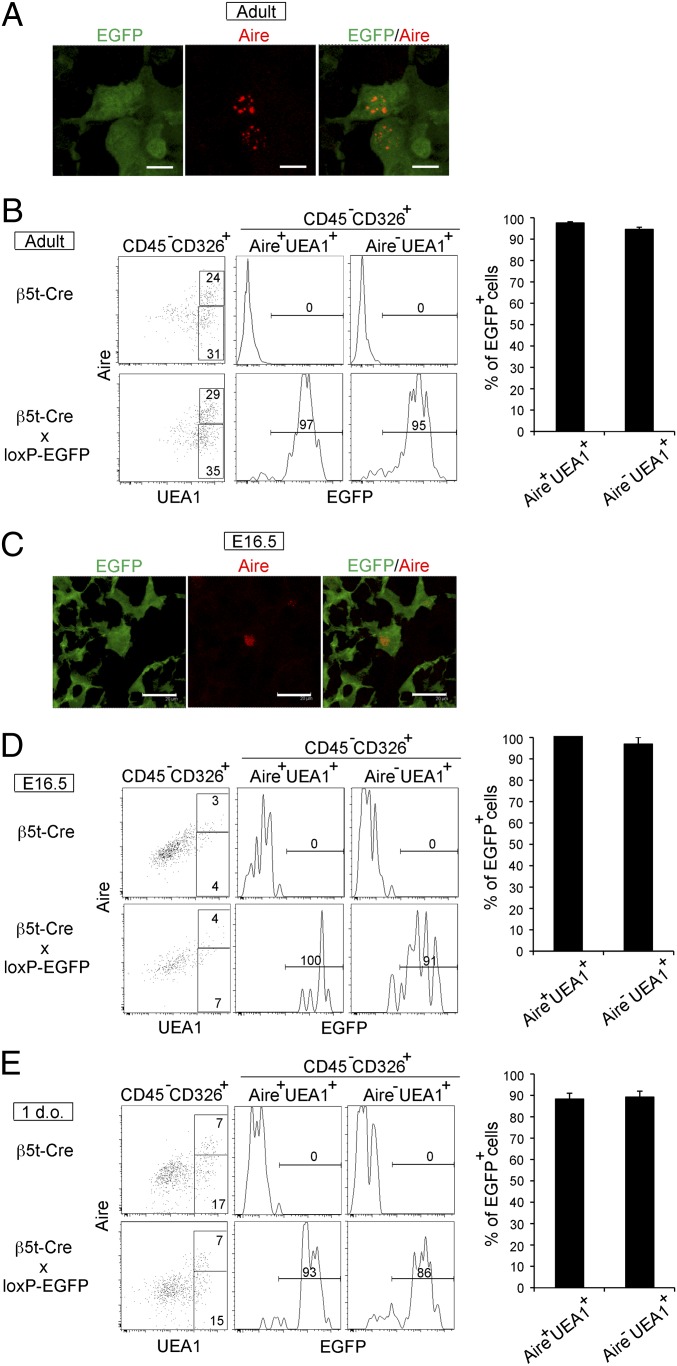
The thymus provides multiple microenvironments that are essential for the development and repertoire selection of T lymphocytes. The thymic cortex induces the generation and positive selection of T lymphocytes, whereas the thymic medulla establishes self-tolerance among the positively selected T lymphocytes. Cortical thymic epithelial cells (cTECs) and medullary TECs (mTECs) constitute the major stromal cells that structurally form and functionally characterize the cortex and the medulla, respectively. cTECs and mTECs are both derived from the endodermal epithelium of the third pharyngeal pouch. However, the molecular and cellular characteristics of the progenitor cells for the distinct TEC lineages are unclear. Here we report the preparation and characterization of mice that express the recombinase Cre instead of β5t, a proteasome subunit that is abundant in cTECs and not detected in other cell types, including mTECs. By crossing β5t-Cre knock-in mice with loxP-dependent GFP reporter mice, we found that β5t-Cre-mediated recombination occurs specifically in TECs but not in any other cell types in the mouse. Surprisingly, in addition to cTECs, β5t-Cre-loxP-mediated GFP expression was detected in almost all mTECs. These results indicate that the majority of mTECs, including autoimmune regulator-expressing mTECs, are derived from β5t-expressing progenitor cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Manley NR, Blackburn CC. A developmental look at thymus organogenesis: Where do the non-hematopoietic cells in the thymus come from? Curr Opin Immunol. 2003;15(2):225–232. - PubMed
-
- Boehm T. Thymus development and function. Curr Opin Immunol. 2008;20(2):178–184. - PubMed
-
- Rodewald HR. Thymus organogenesis. Annu Rev Immunol. 2008;26:355–388. - PubMed
-
- Klein L, Hinterberger M, Wirnsberger G, Kyewski B. Antigen presentation in the thymus for positive selection and central tolerance induction. Nat Rev Immunol. 2009;9(12):833–844. - PubMed
-
- Anderson G, Takahama Y. Thymic epithelial cells: Working class heroes for T cell development and repertoire selection. Trends Immunol. 2012;33(6):256–263. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
