

Impact of plant shoot architecture on leaf cooling: a coupled heat and mass transfer model
- PMID: 23720538
- PMCID: PMC4043166
- DOI: 10.1098/rsif.2013.0326
Impact of plant shoot architecture on leaf cooling: a coupled heat and mass transfer model
Abstract
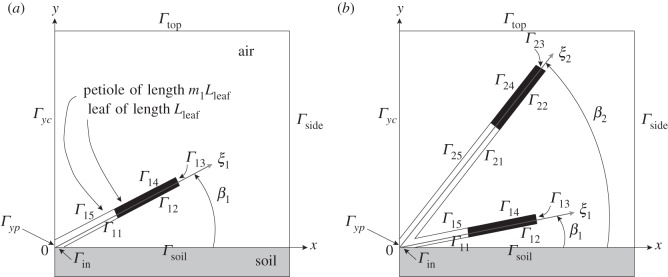
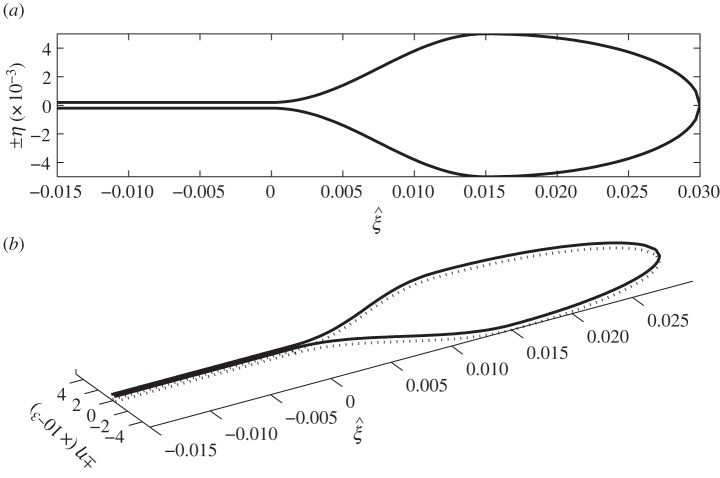
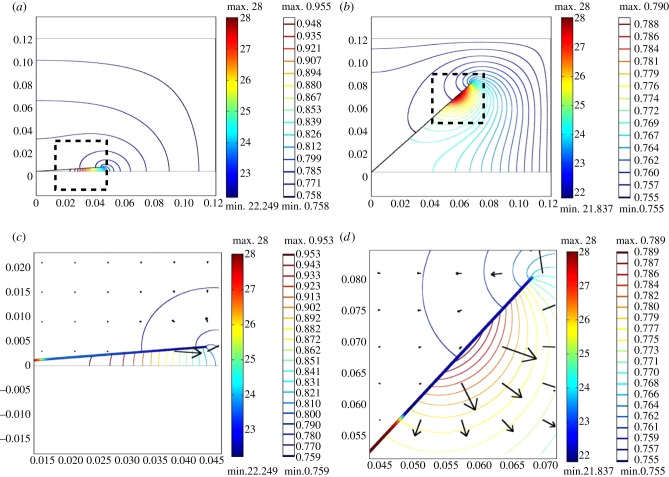
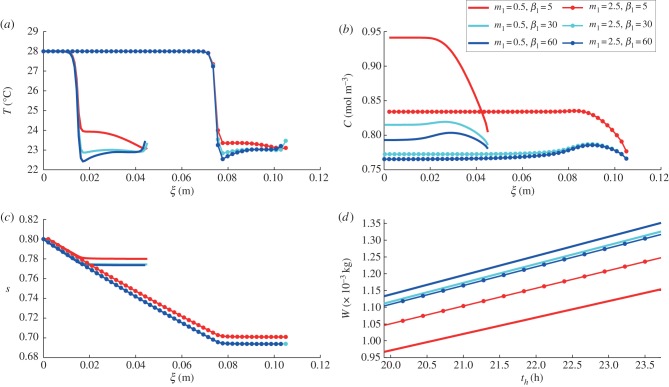
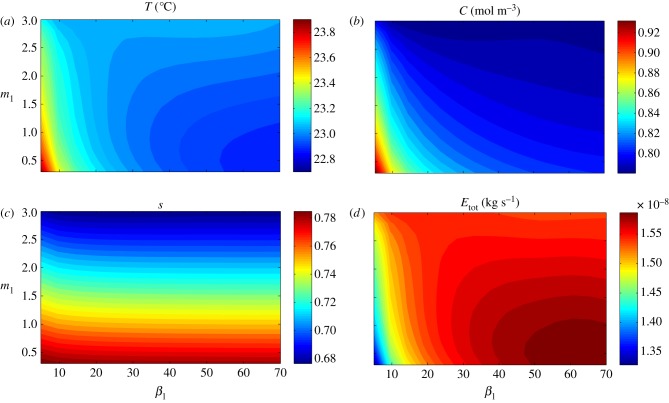
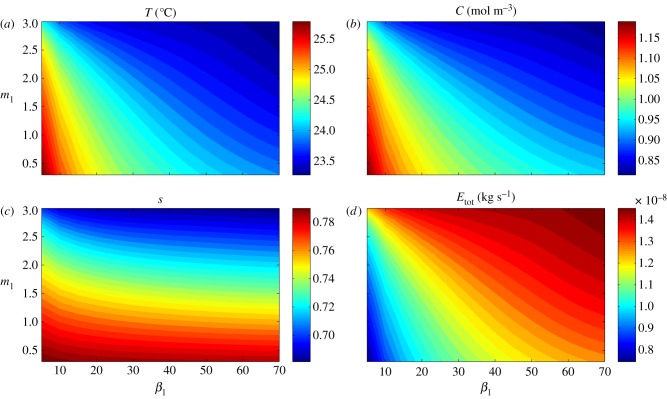
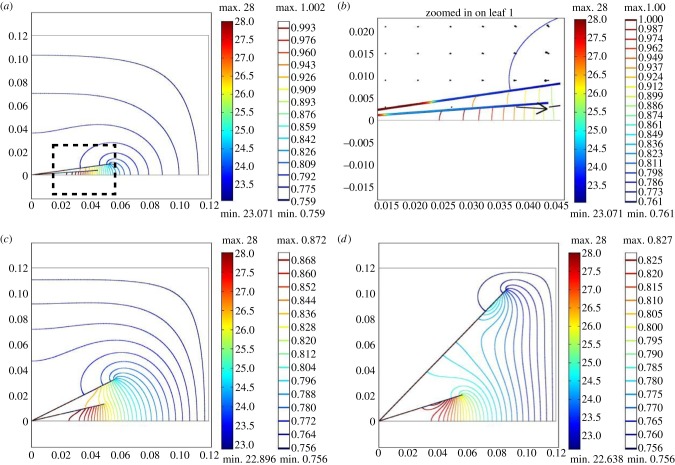
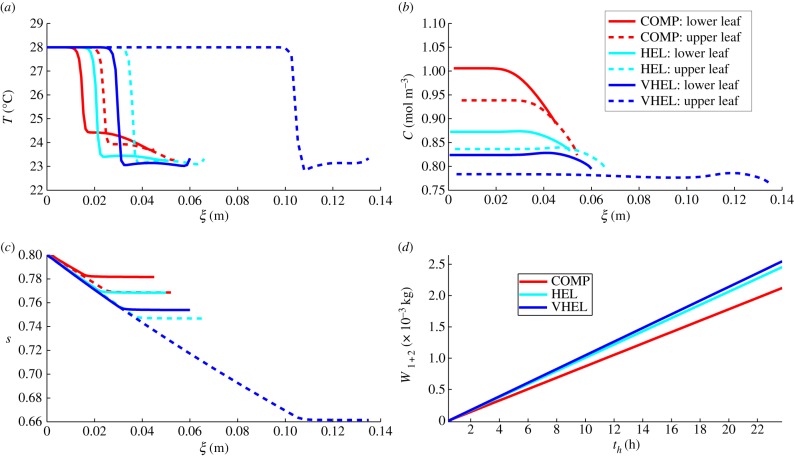
Plants display a range of striking architectural adaptations when grown at elevated temperatures. In the model plant Arabidopsis thaliana, these include elongation of petioles, and increased petiole and leaf angles from the soil surface. The potential physiological significance of these architectural changes remains speculative. We address this issue computationally by formulating a mathematical model and performing numerical simulations, testing the hypothesis that elongated and elevated plant configurations may reflect a leaf-cooling strategy. This sets in place a new basic model of plant water use and interaction with the surrounding air, which couples heat and mass transfer within a plant to water vapour diffusion in the air, using a transpiration term that depends on saturation, temperature and vapour concentration. A two-dimensional, multi-petiole shoot geometry is considered, with added leaf-blade shape detail. Our simulations show that increased petiole length and angle generally result in enhanced transpiration rates and reduced leaf temperatures in well-watered conditions. Furthermore, our computations also reveal plant configurations for which elongation may result in decreased transpiration rate owing to decreased leaf liquid saturation. We offer further qualitative and quantitative insights into the role of architectural parameters as key determinants of leaf-cooling capacity.
Keywords: computational partial differential equations; leaf hyponasty; mathematical modelling; petiole elongation; plant architecture; transpiration.
Figures

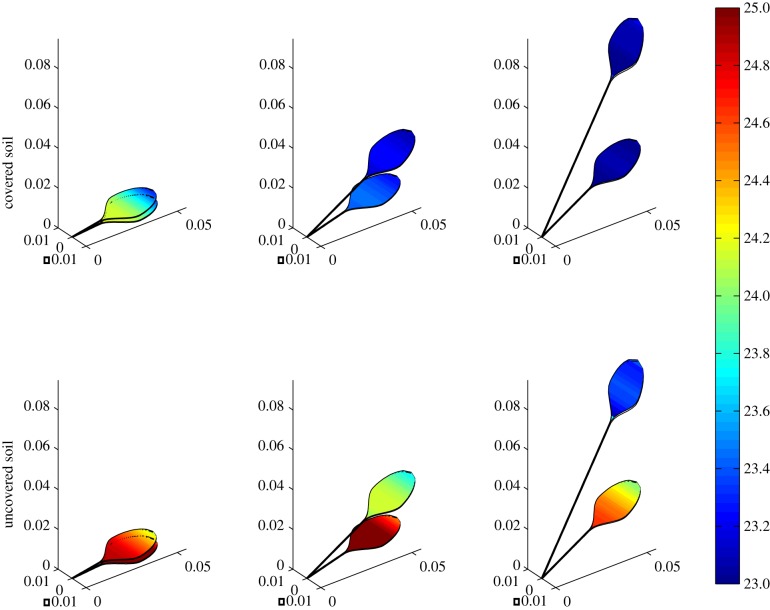
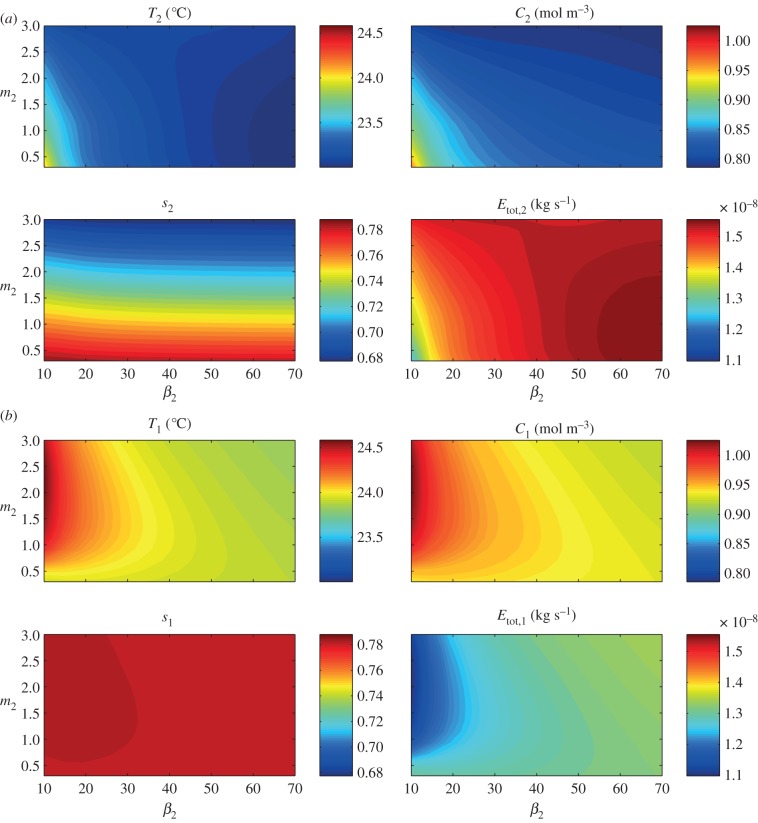
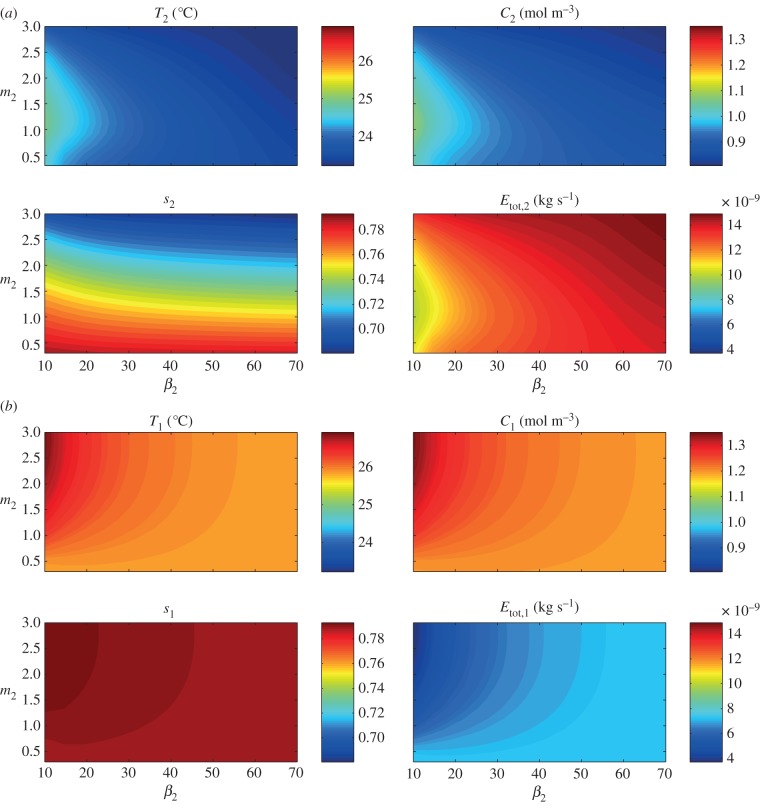
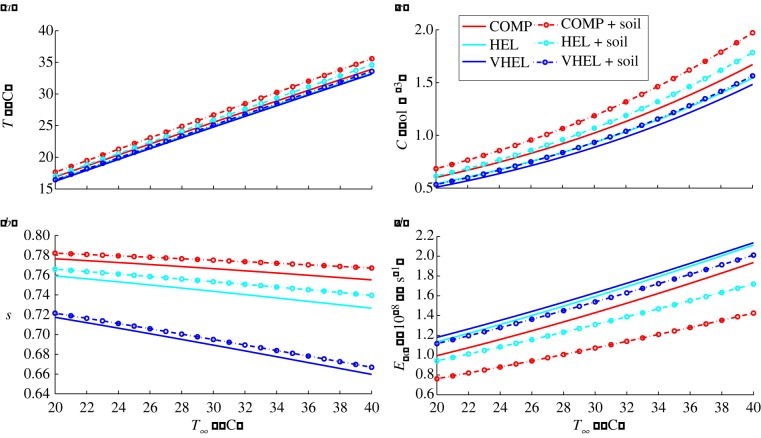
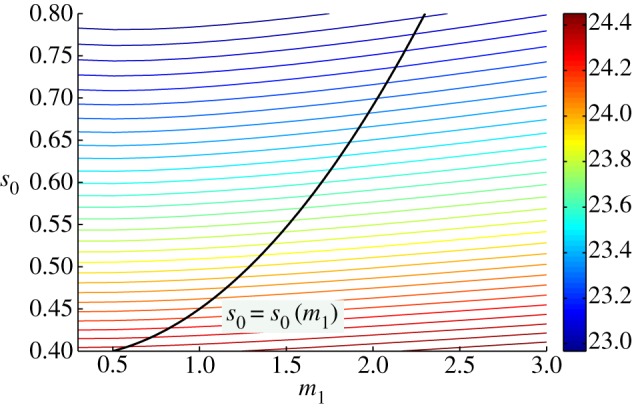
References
-
- Koini MA, Alvey L, Allen T, Tilley CA, Harberd NP, Whitelam GC, Franklin KA. 2009. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol. 19, 408–413 10.1016/j.cub.2009.01.046 (doi:10.1016/j.cub.2009.01.046) - DOI - PubMed
-
- Crawford AJ, McLachlan DH, Hetherington AM, Franklin KA. 2012. High temperature exposure increases plant cooling capacity. Curr. Biol. 22, R396–R397 10.1016/j.cub.2012.03.044 (doi:10.1016/j.cub.2012.03.044) - DOI - PubMed
-
- Gray WM, Östin A, Sandberg G, Romano CP, Estelle M. 1998. High temperature promotes auxin-mediated hypocotyl elongation in Arabidopsis. Proc. Natl Acad. Sci. USA 95, 7197–7202 10.1073/pnas.95.12.7197 (doi:10.1073/pnas.95.12.7197) - DOI - PMC - PubMed
-
- Stavang JA, Gallego-Bartolome J, Gomez M, Yoshida S, Asami T, Olsen JE, Garca-Martnez JL, Alabadi D, Blazquez MA. 2009. Hormonal regulation of temperature-induced growth in Arabidopsis. Plant J. 60, 589–601 10.1111/j.1365-313X.2009.03983.x (doi:10.1111/j.1365-313X.2009.03983.x) - DOI - PubMed
-
- Franklin KA, et al. 2011. Phytochrome interacting factor 4 regulates auxin biosynthesis at high temperature. Proc. Natl Acad. Sci. USA 108, 20 231–20 235 10.1073/pnas.1110682108 (doi:10.1073/pnas.1110682108) - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources