

Dominant protein interactions that influence the pathogenesis of conformational diseases
- PMID: 23722904
- PMCID: PMC3696544
- DOI: 10.1172/JCI67260
Dominant protein interactions that influence the pathogenesis of conformational diseases
Abstract
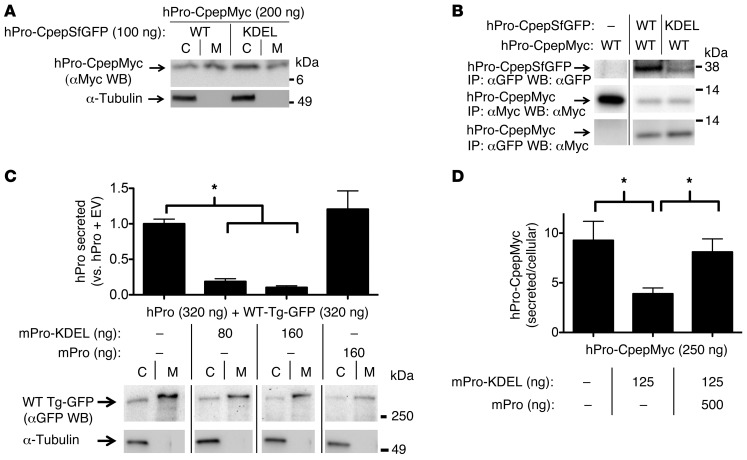
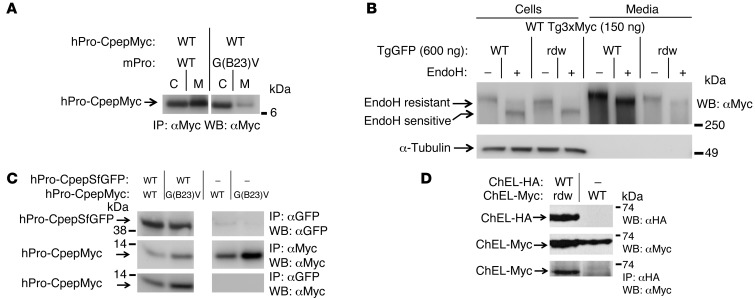
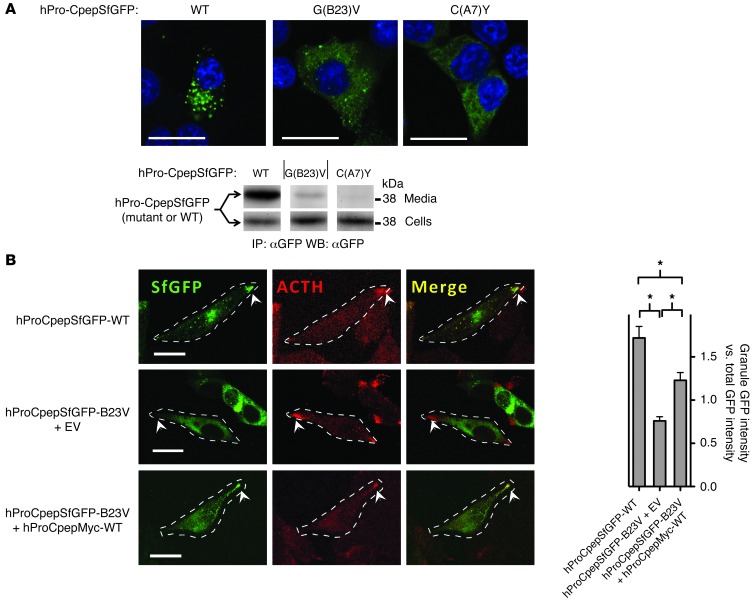
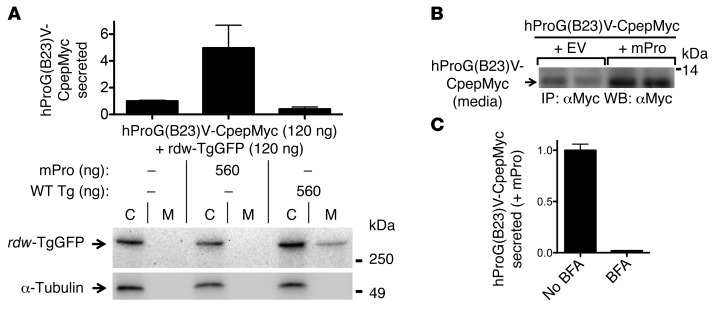
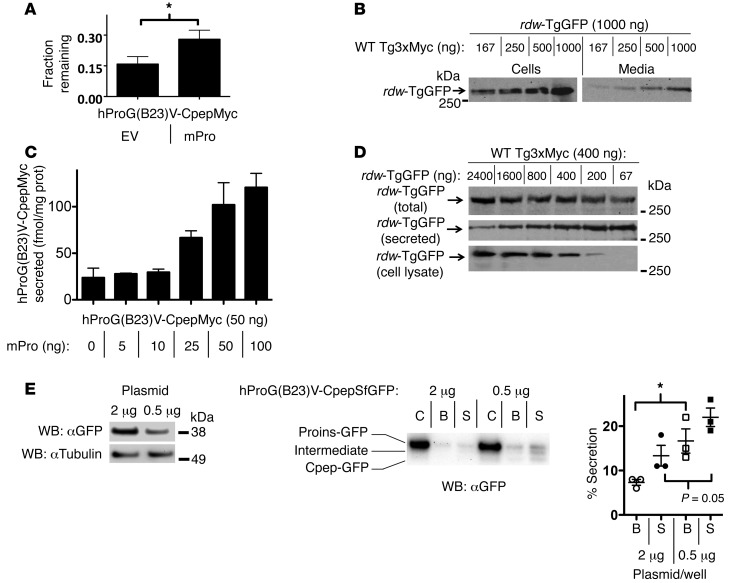
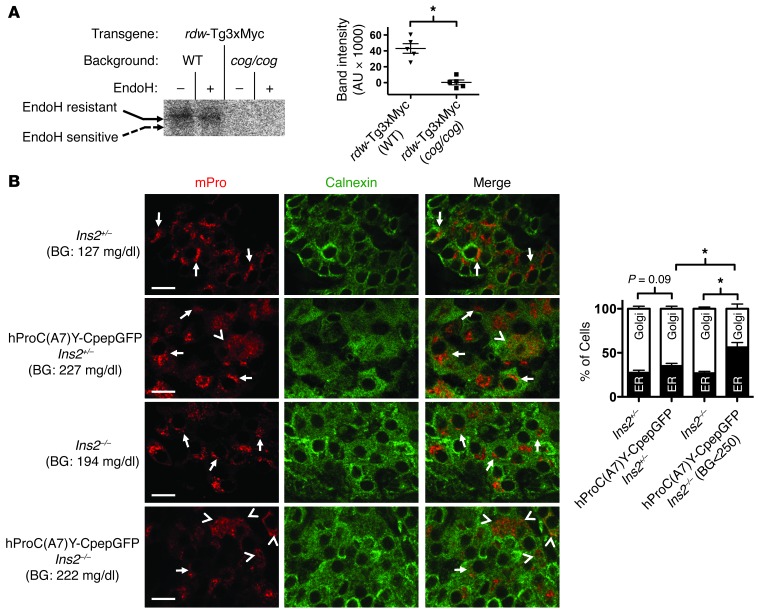
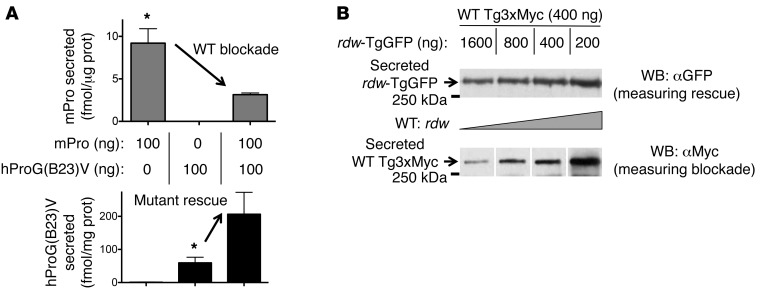
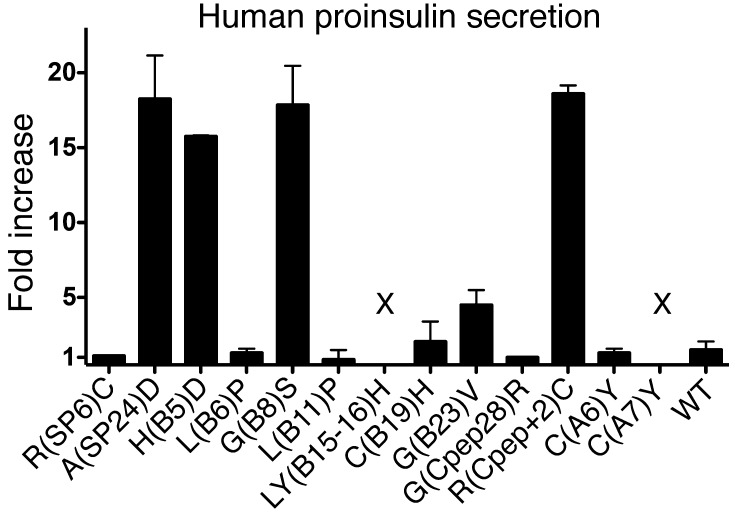
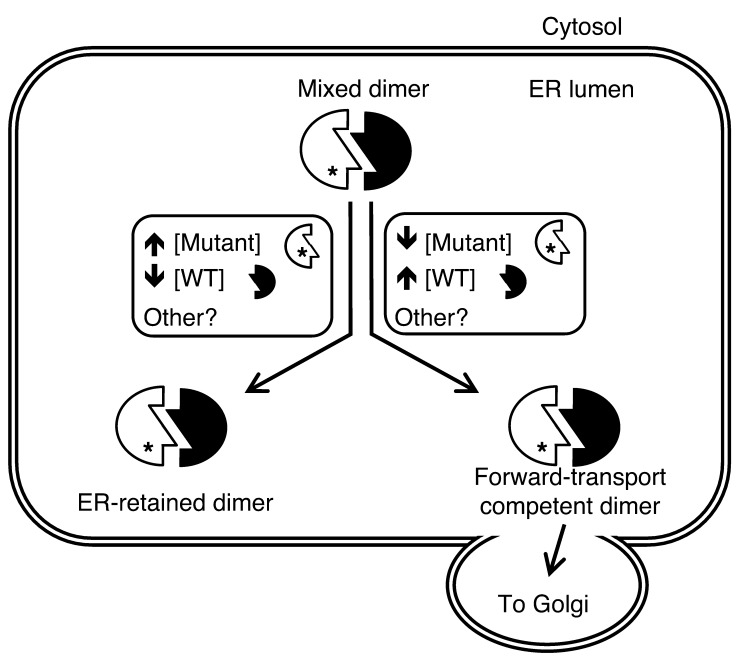
Misfolding of exportable proteins can trigger endocrinopathies. For example, misfolding of insulin can result in autosomal dominant mutant INS gene-induced diabetes of youth, and misfolding of thyroglobulin can result in autosomal recessive congenital hypothyroidism with deficient thyroglobulin. Both proinsulin and thyroglobulin normally form homodimers; the mutant versions of both proteins misfold in the ER, triggering ER stress, and, in both cases, heterozygosity creates potential for cross-dimerization between mutant and WT gene products. Here, we investigated these two ER-retained mutant secretory proteins and the selectivity of their interactions with their respective WT counterparts. In both cases and in animal models of these diseases, we found that conditions favoring an increased stoichiometry of mutant gene product dominantly inhibited export of the WT partner, while increased relative level of the WT gene product helped to rescue secretion of the mutant partner. Surprisingly, the bidirectional consequences of secretory blockade and rescue occur simultaneously in the same cells. Thus, in the context of heterozygosity, expression level and stability of WT subunits may be a critical factor influencing the effect of protein misfolding on clinical phenotype. These results offer new insight into dominant as well as recessive inheritance of conformational diseases and offer opportunities for the development of new therapies.
Figures
Similar articles
-
Proinsulin misfolding and diabetes: mutant INS gene-induced diabetes of youth.Trends Endocrinol Metab. 2010 Nov;21(11):652-9. doi: 10.1016/j.tem.2010.07.001. Epub 2010 Aug 18. Trends Endocrinol Metab. 2010. PMID: 20724178 Free PMC article. Review.
-
Endoplasmic reticulum oxidoreductin-1α (Ero1α) improves folding and secretion of mutant proinsulin and limits mutant proinsulin-induced endoplasmic reticulum stress.J Biol Chem. 2013 Oct 25;288(43):31010-8. doi: 10.1074/jbc.M113.510065. Epub 2013 Sep 10. J Biol Chem. 2013. PMID: 24022479 Free PMC article.
-
Biological behaviors of mutant proinsulin contribute to the phenotypic spectrum of diabetes associated with insulin gene mutations.Mol Cell Endocrinol. 2020 Dec 1;518:111025. doi: 10.1016/j.mce.2020.111025. Epub 2020 Sep 8. Mol Cell Endocrinol. 2020. PMID: 32916194 Free PMC article.
-
Diabetes mellitus due to the toxic misfolding of proinsulin variants.FEBS Lett. 2013 Jun 27;587(13):1942-50. doi: 10.1016/j.febslet.2013.04.044. Epub 2013 May 10. FEBS Lett. 2013. PMID: 23669362 Free PMC article. Review.
-
Role of Proinsulin Self-Association in Mutant INS Gene-Induced Diabetes of Youth.Diabetes. 2020 May;69(5):954-964. doi: 10.2337/db19-1106. Epub 2020 Mar 5. Diabetes. 2020. PMID: 32139596 Free PMC article.
Cited by
-
INS-gene mutations: from genetics and beta cell biology to clinical disease.Mol Aspects Med. 2015 Apr;42:3-18. doi: 10.1016/j.mam.2014.12.001. Epub 2014 Dec 24. Mol Aspects Med. 2015. PMID: 25542748 Free PMC article. Review.
-
Lessons from animal models of endocrine disorders caused by defects of protein folding in the secretory pathway.Mol Cell Endocrinol. 2020 Jan 1;499:110613. doi: 10.1016/j.mce.2019.110613. Epub 2019 Oct 9. Mol Cell Endocrinol. 2020. PMID: 31605742 Free PMC article. Review.
-
Defective transport of the obesity mutant PC1/3 N222D contributes to loss of function.Endocrinology. 2014 Jul;155(7):2391-401. doi: 10.1210/en.2013-1985. Epub 2014 May 14. Endocrinology. 2014. PMID: 24828610 Free PMC article.
-
Misfolded proinsulin impairs processing of precursor of insulin receptor and insulin signaling in β cells.FASEB J. 2019 Oct;33(10):11338-11348. doi: 10.1096/fj.201900442R. Epub 2019 Aug 1. FASEB J. 2019. PMID: 31311313 Free PMC article.
-
Surveying the floodgates: estimating protein flux into the endoplasmic reticulum lumen in Saccharomyces cerevisiae.Front Physiol. 2014 Nov 13;5:444. doi: 10.3389/fphys.2014.00444. eCollection 2014. Front Physiol. 2014. PMID: 25431559 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- T32GM007863/GM/NIGMS NIH HHS/United States
- T32GM008322/GM/NIGMS NIH HHS/United States
- F30 DK095504/DK/NIDDK NIH HHS/United States
- R01DK40344/DK/NIDDK NIH HHS/United States
- F30DK095504/DK/NIDDK NIH HHS/United States
- P30DK034933/DK/NIDDK NIH HHS/United States
- R01 DK048280/DK/NIDDK NIH HHS/United States
- T32 HD007505/HD/NICHD NIH HHS/United States
- P30DK020572/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- R01DK48280/DK/NIDDK NIH HHS/United States
- T32HD007505/HD/NICHD NIH HHS/United States
- T32 GM007863/GM/NIGMS NIH HHS/United States
- T32 GM008322/GM/NIGMS NIH HHS/United States
- R01 DK040344/DK/NIDDK NIH HHS/United States
- P30 DK034933/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
