

Disentangling the origins of cultivated sweet potato (Ipomoea batatas (L.) Lam.)
- PMID: 23723970
- PMCID: PMC3664560
- DOI: 10.1371/journal.pone.0062707
Disentangling the origins of cultivated sweet potato (Ipomoea batatas (L.) Lam.)
Abstract
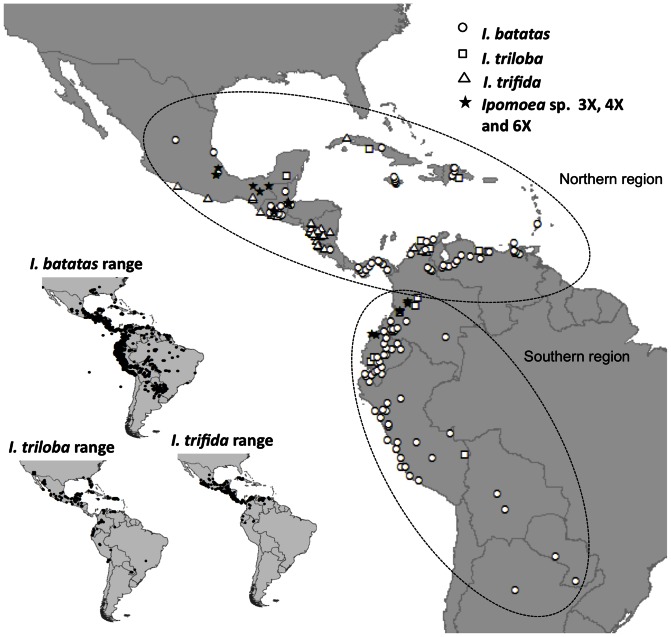
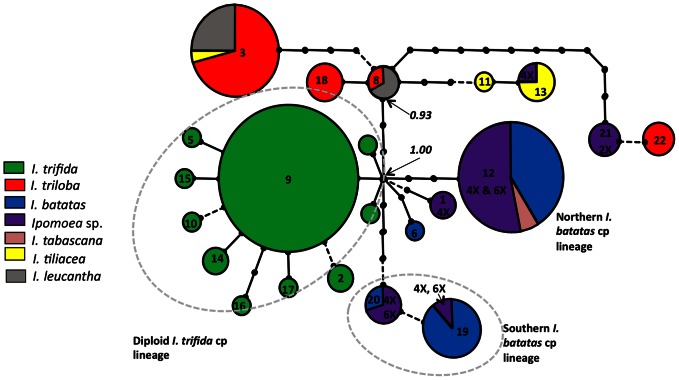
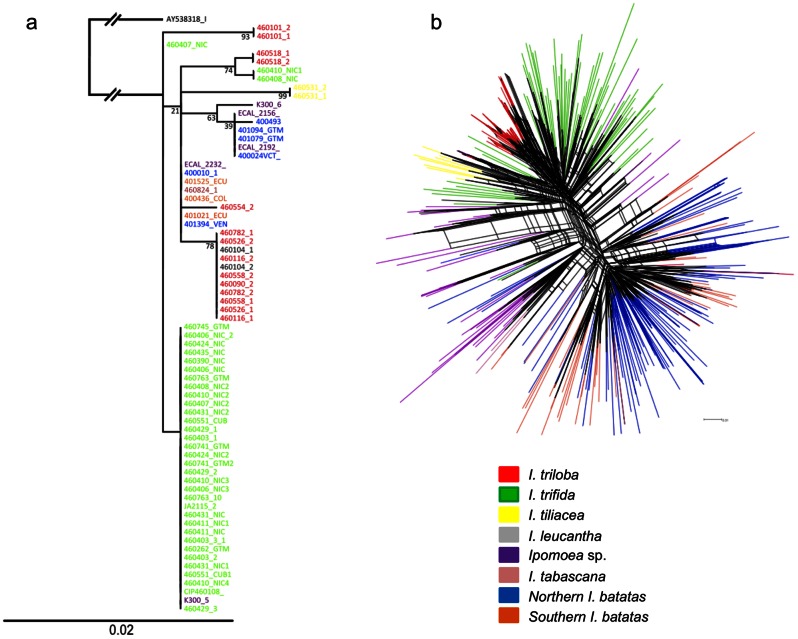
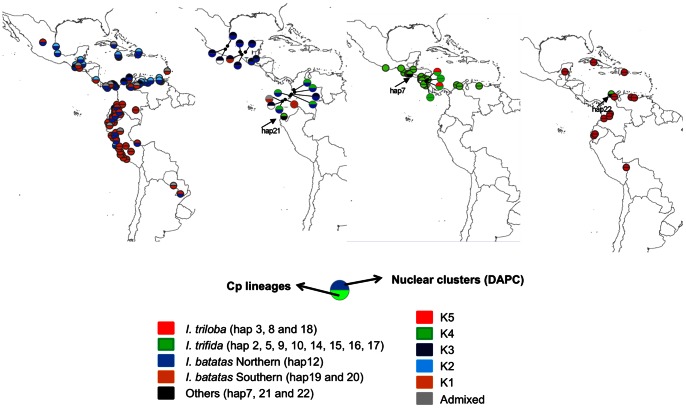
Sweet potato (Ipomoea batatas (L.) Lam., Convolvulaceae) counts among the most widely cultivated staple crops worldwide, yet the origins of its domestication remain unclear. This hexaploid species could have had either an autopolyploid origin, from the diploid I. trifida, or an allopolyploid origin, involving genomes of I. trifida and I. triloba. We generated molecular genetic data for a broad sample of cultivated sweet potatoes and its diploid and polyploid wild relatives, for noncoding chloroplast and nuclear ITS sequences, and nuclear SSRs. Our data did not support an allopolyploid origin for I. batatas, nor any contribution of I. triloba in the genome of domesticated sweet potato. I. trifida and I. batatas are closely related although they do not share haplotypes. Our data support an autopolyploid origin of sweet potato from the ancestor it shares with I. trifida, which might be similar to currently observed tetraploid wild Ipomoea accessions. Two I. batatas chloroplast lineages were identified. They show more divergence with each other than either does with I. trifida. We thus propose that cultivated I. batatas have multiple origins, and evolved from at least two distinct autopolyploidization events in polymorphic wild populations of a single progenitor species. Secondary contact between sweet potatoes domesticated in Central America and in South America, from differentiated wild I. batatas populations, would have led to the introgression of chloroplast haplotypes of each lineage into nuclear backgrounds of the other, and to a reduced divergence between nuclear gene pools as compared with chloroplast haplotypes.
Conflict of interest statement
Figures

References
-
- Otto SP (2007) The evolutionary consequences of polyploidy. Cell 131: 452–462. - PubMed
-
- Purugganan MD, Fuller DQ (2009) The nature of selection during plant domestication. Nature 457: 843–848. - PubMed
-
- Zohary D (2004) Unconscious selection and the evolution of domesticated plants. Econ Bot 58: 5–10.
-
- McKey D, Elias M, Pujol B, Duputié A (2010) The evolutionary ecology of clonally propagated domesticated plants. New Phytol 186: 318–332. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
