

Spatial distribution of prominin-1 (CD133)-positive cells within germinative zones of the vertebrate brain
- PMID: 23723983
- PMCID: PMC3664558
- DOI: 10.1371/journal.pone.0063457
Spatial distribution of prominin-1 (CD133)-positive cells within germinative zones of the vertebrate brain
Abstract
Background: In mammals, embryonic neural progenitors as well as adult neural stem cells can be prospectively isolated based on the cell surface expression of prominin-1 (CD133), a plasma membrane glycoprotein. In contrast, characterization of neural progenitors in non-mammalian vertebrates endowed with significant constitutive neurogenesis and inherent self-repair ability is hampered by the lack of suitable cell surface markers. Here, we have investigated whether prominin-1-orthologues of the major non-mammalian vertebrate model organisms show any degree of conservation as for their association with neurogenic geminative zones within the central nervous system (CNS) as they do in mammals or associated with activated neural progenitors during provoked neurogenesis in the regenerating CNS.
Methods: We have recently identified prominin-1 orthologues from zebrafish, axolotl and chicken. The spatial distribution of prominin-1-positive cells--in comparison to those of mice--was mapped in the intact brain in these organisms by non-radioactive in situ hybridization combined with detection of proliferating neural progenitors, marked either by proliferating cell nuclear antigen or 5-bromo-deoxyuridine. Furthermore, distribution of prominin-1 transcripts was investigated in the regenerating spinal cord of injured axolotl.
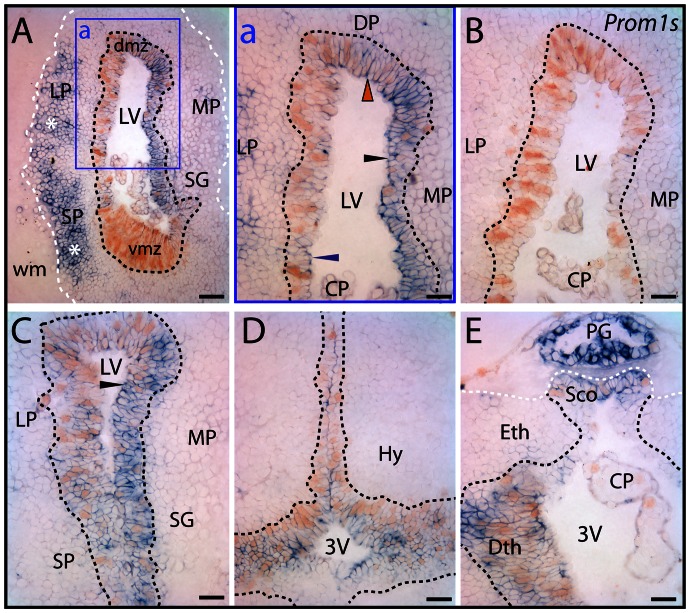
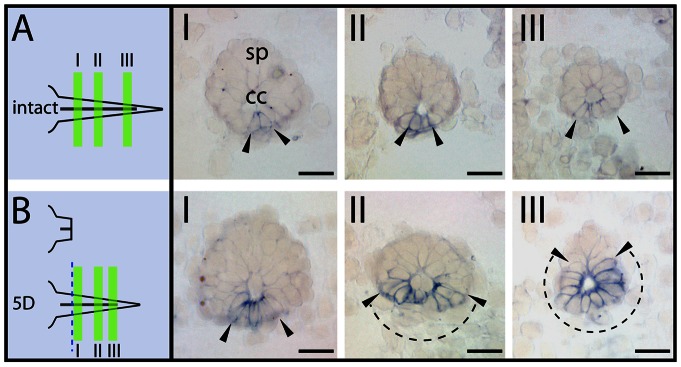
Results: Remarkably, a conserved association of prominin-1 with germinative zones of the CNS was uncovered as manifested in a significant co-localization with cell proliferation markers during normal constitutive neurogenesis in all species investigated. Moreover, an enhanced expression of prominin-1 became evident associated with provoked, compensatory neurogenesis during the epimorphic regeneration of the axolotl spinal cord. Interestingly, significant prominin-1-expressing cell populations were also detected at distinct extraventricular (parenchymal) locations in the CNS of all vertebrate species being suggestive of further, non-neurogenic neural function(s).
Conclusion/interpretation: Collectively, our work provides the first data set describing a comparative analysis of prominin-1-positive progenitor cells across species establishing a framework for further functional characterization in the context of regeneration.
Conflict of interest statement
Figures
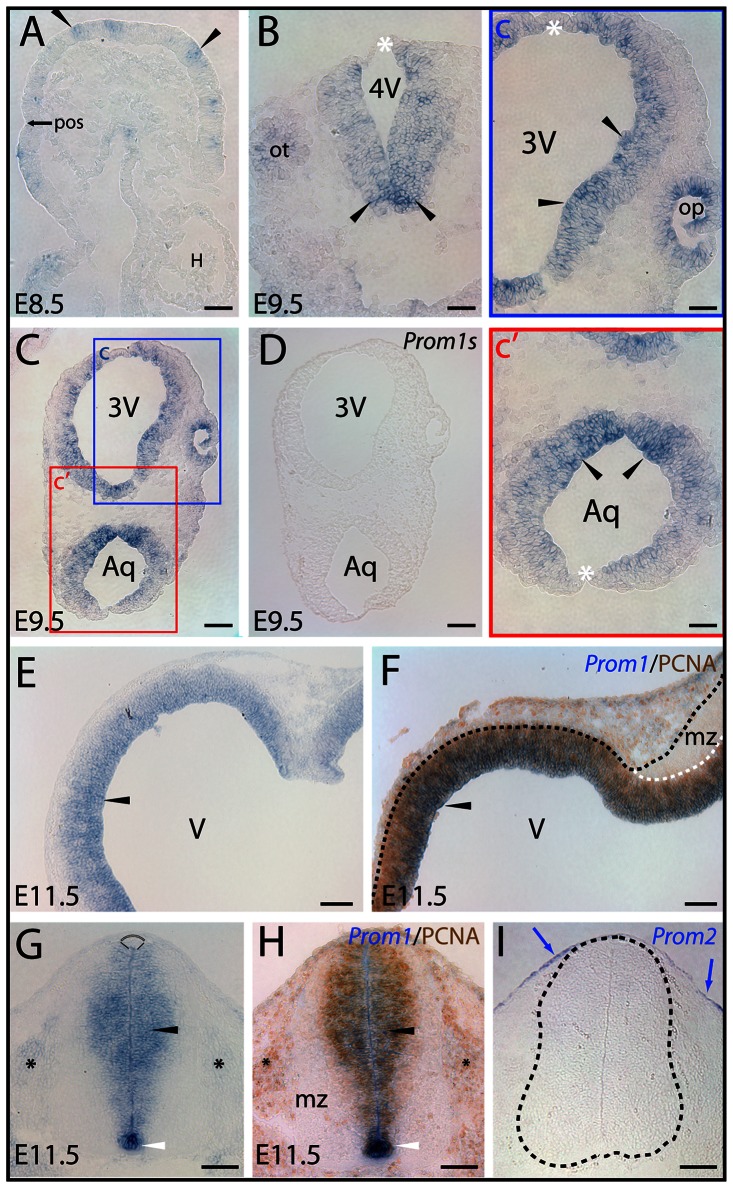
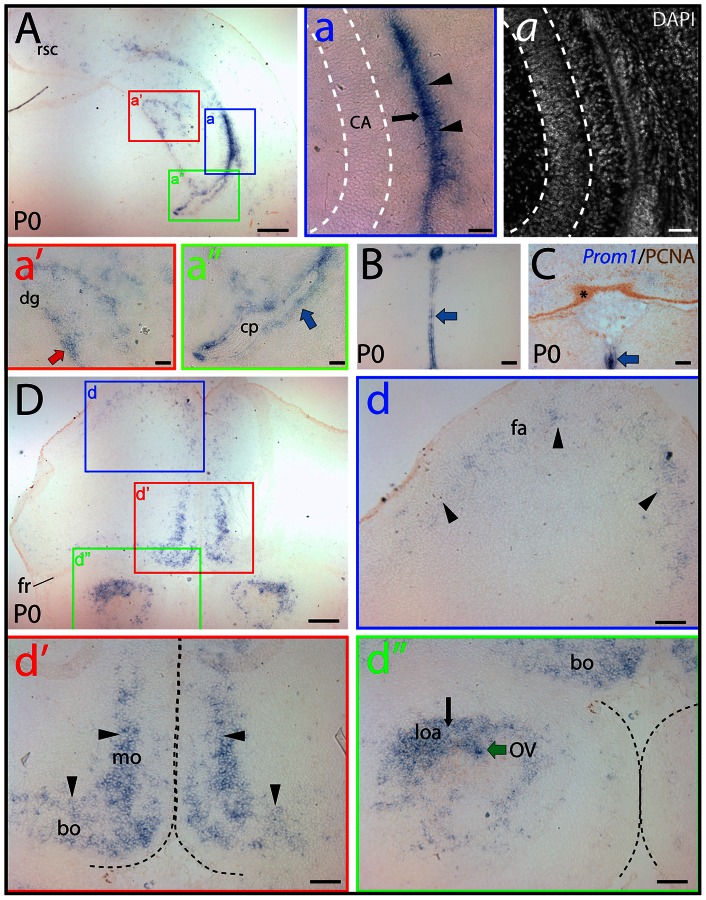
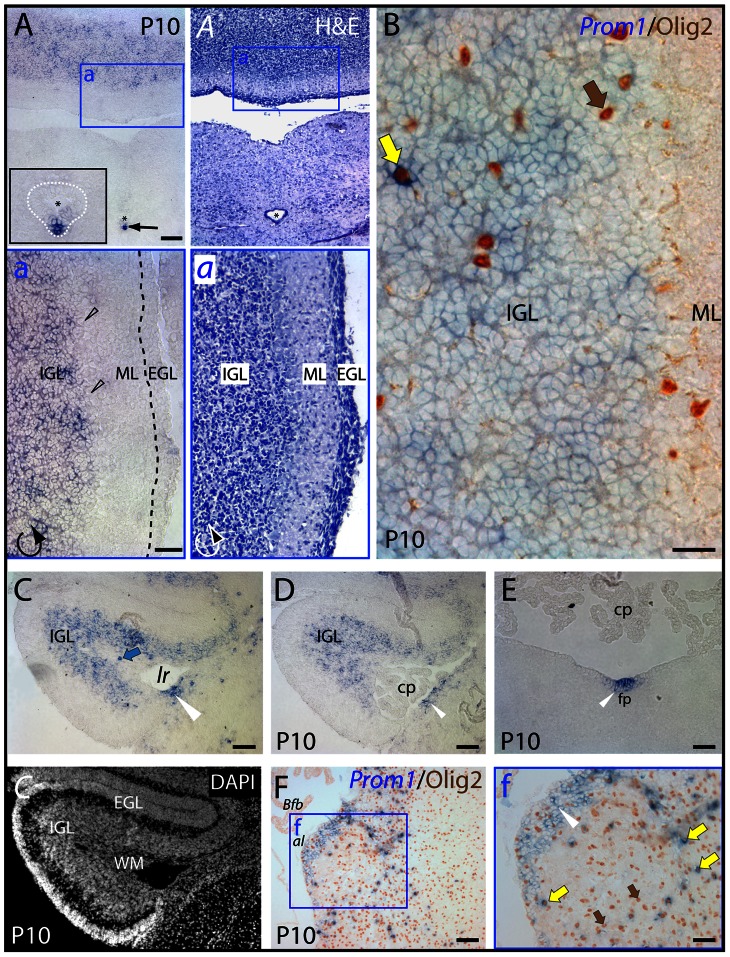
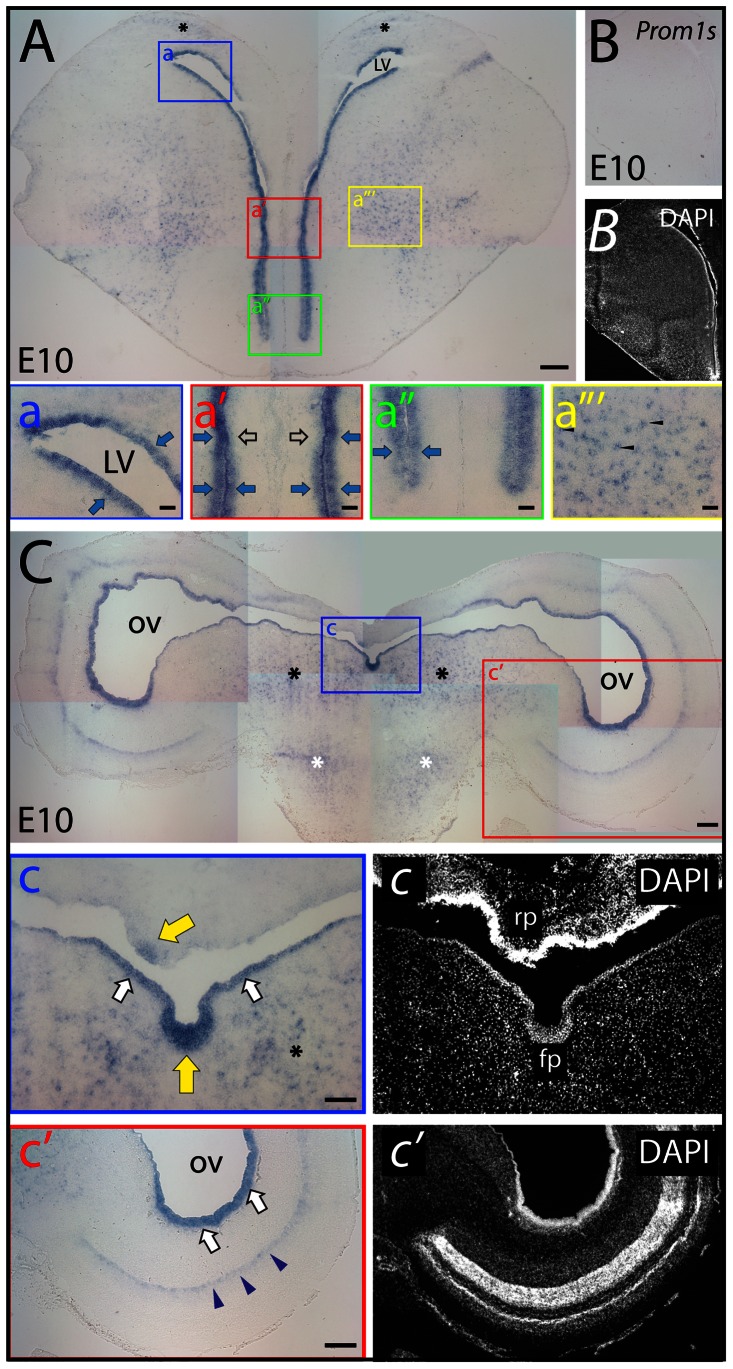
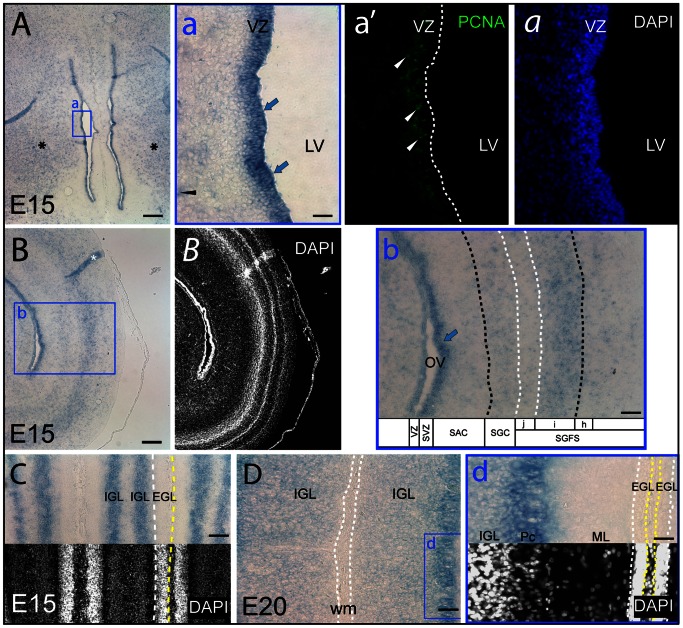
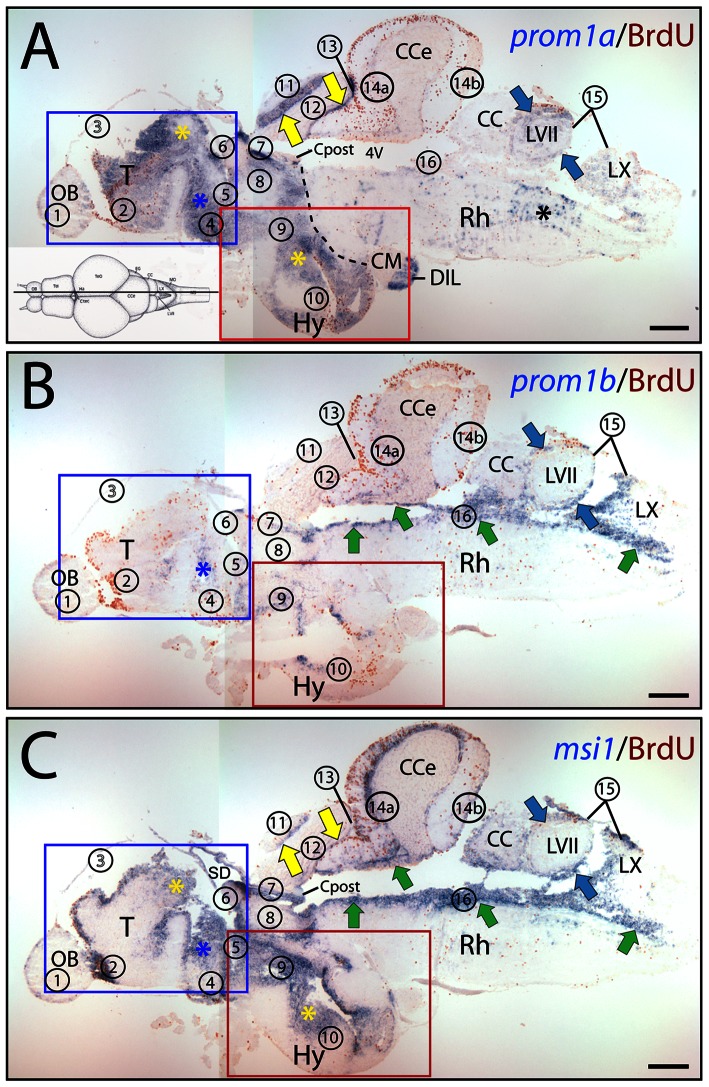
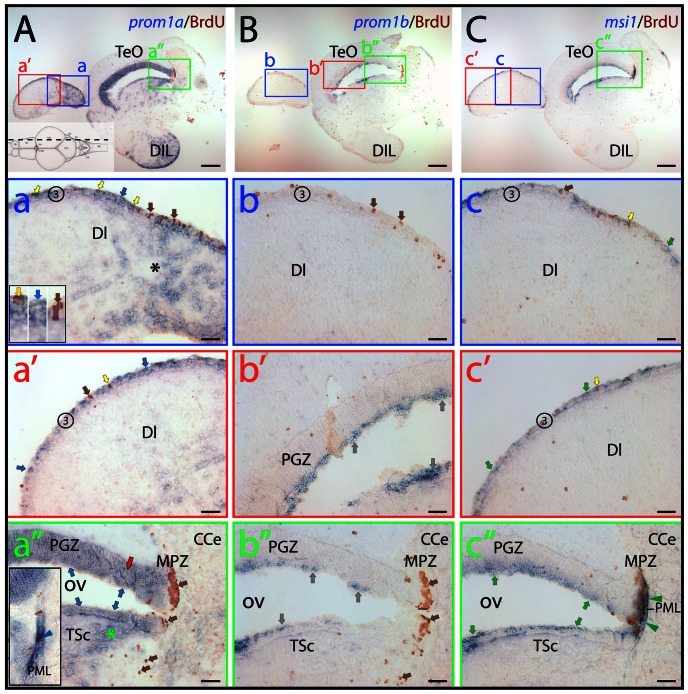
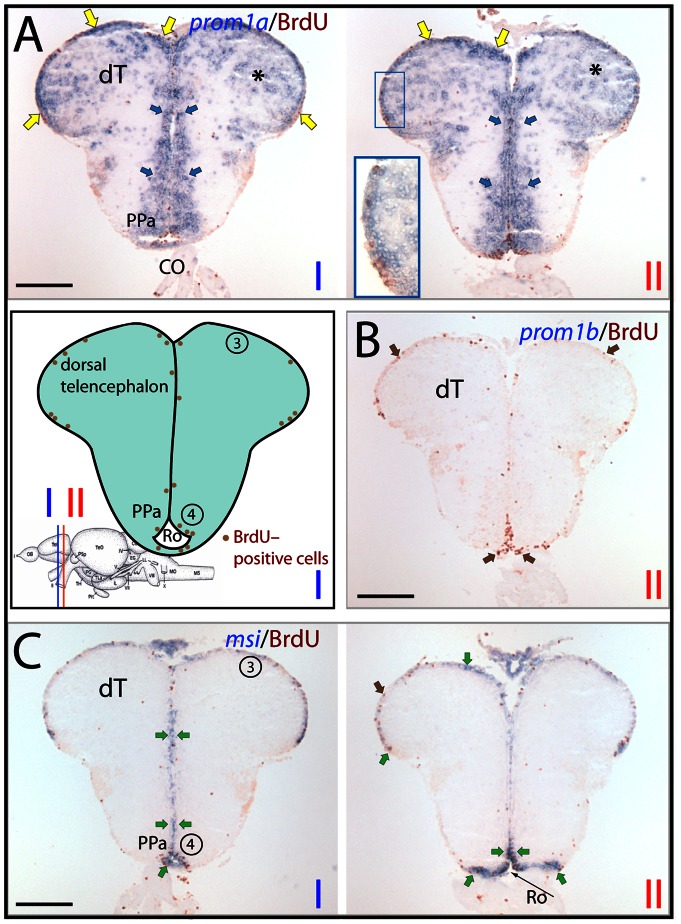
References
-
- Doetsch F, Caillé I, Lim DA, García-Verdugo JM, Alvarez-Buylla A (1999) Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97: 703–16. - PubMed
-
- Alvarez-Buylla A, Seri B, Doetsch F (2002) Identification of neural stem cells in the adult vertebrate brain. Brain Res Bull 57: 751–58. - PubMed
-
- Götz M, Huttner WB (2005) The cell biology of neurogenesis. Nat Rev Mol Cell Biol 6: 777–88. - PubMed
-
- Kee N, Teixeira CM, Wang AH, Frankland PW (2007) Preferential incorporation of adult-generated granule cells into spatial memory networks in the dentate gyrus. Nat Neurosci 10: 355–62. - PubMed
-
- Imayoshi I, Sakamoto M, Ohtsuka T, Takao K, Miyakawa T, et al. (2008) Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nat Neurosci 11: 1153–61. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
