

The organization and physiology of the auditory thalamus and its role in processing acoustic features important for speech perception
- PMID: 23725661
- PMCID: PMC3707394
- DOI: 10.1016/j.bandl.2013.03.003
The organization and physiology of the auditory thalamus and its role in processing acoustic features important for speech perception
Abstract
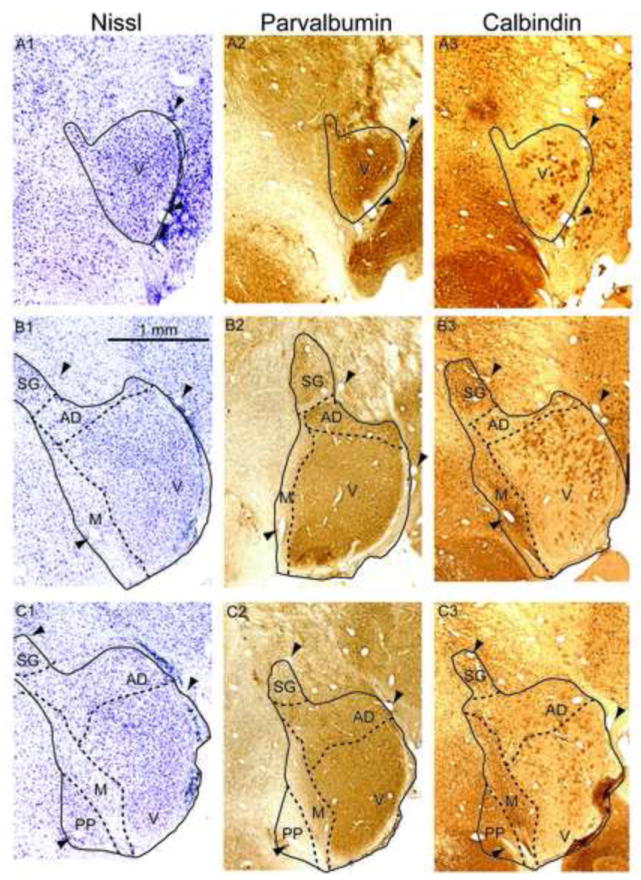
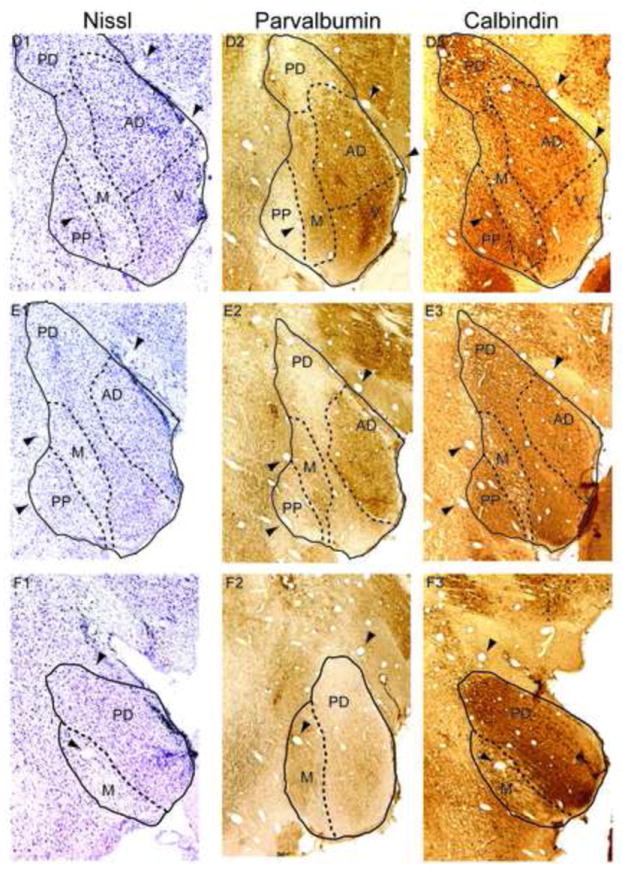
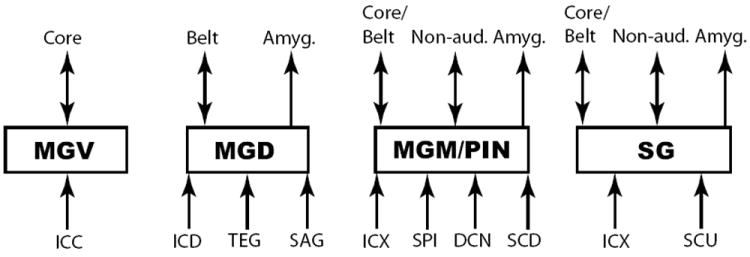
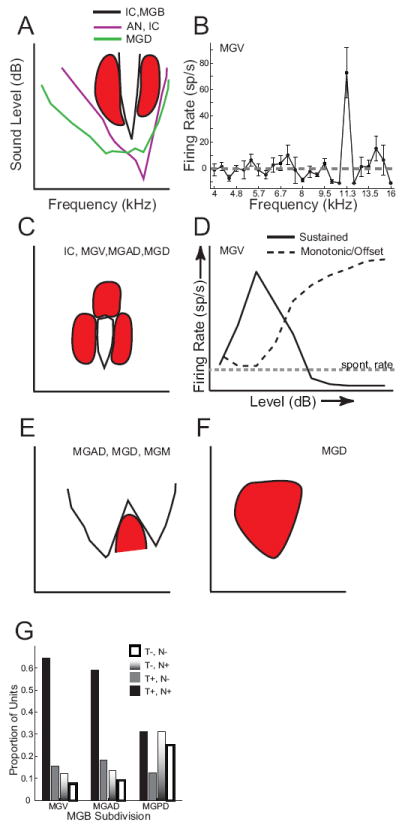
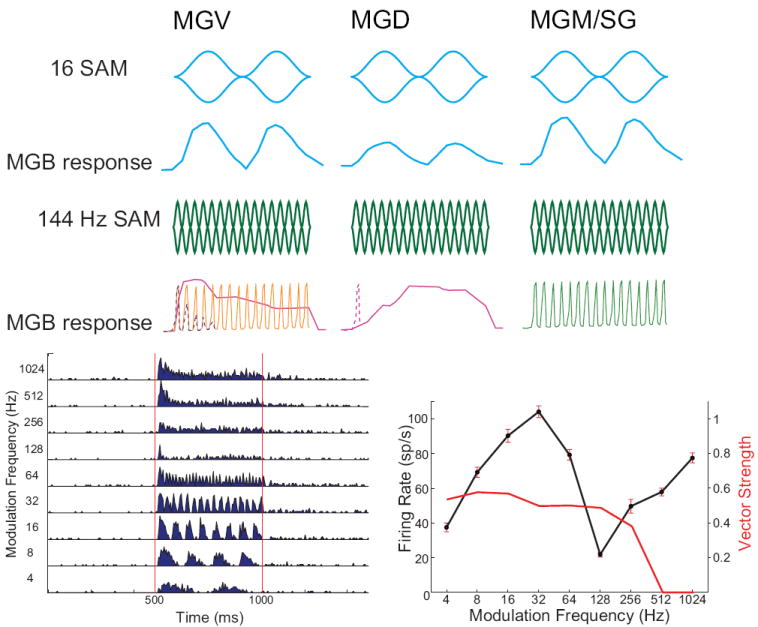
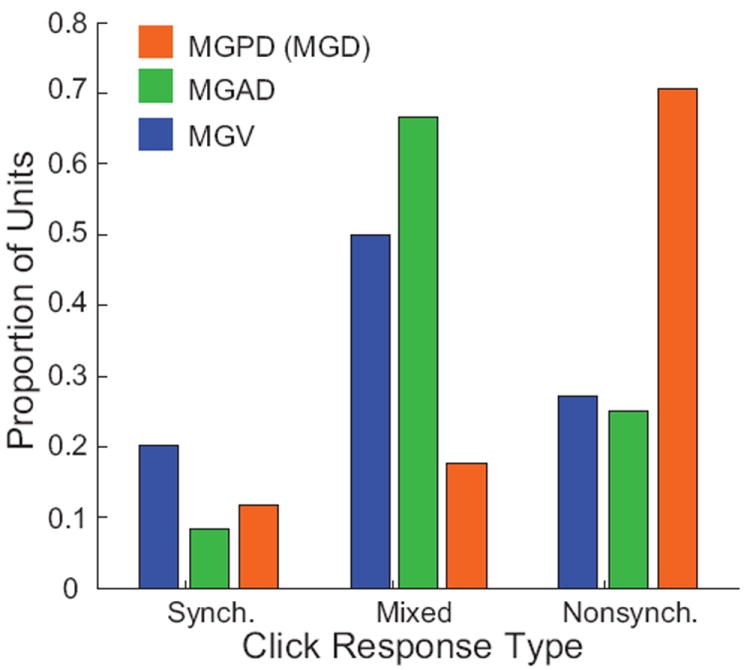
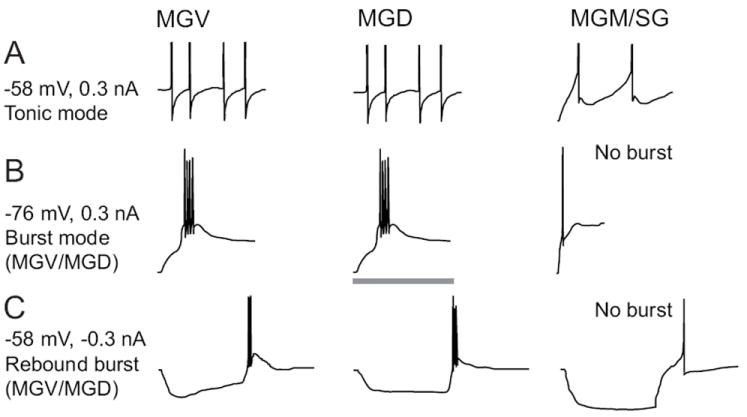
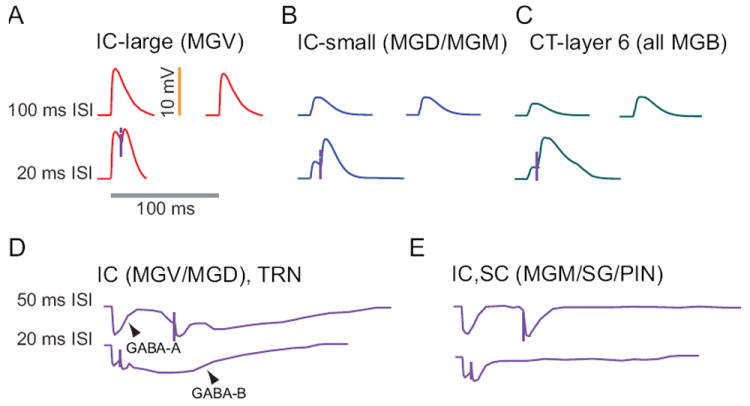
The auditory thalamus, or medial geniculate body (MGB), is the primary sensory input to auditory cortex. Therefore, it plays a critical role in the complex auditory processing necessary for robust speech perception. This review will describe the functional organization of the thalamus as it relates to processing acoustic features important for speech perception, focusing on thalamic nuclei that relate to auditory representations of language sounds. The MGB can be divided into three main subdivisions, the ventral, dorsal, and medial subdivisions, each with different connectivity, auditory response properties, neuronal properties, and synaptic properties. Together, the MGB subdivisions actively and dynamically shape complex auditory processing and form ongoing communication loops with auditory cortex and subcortical structures.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
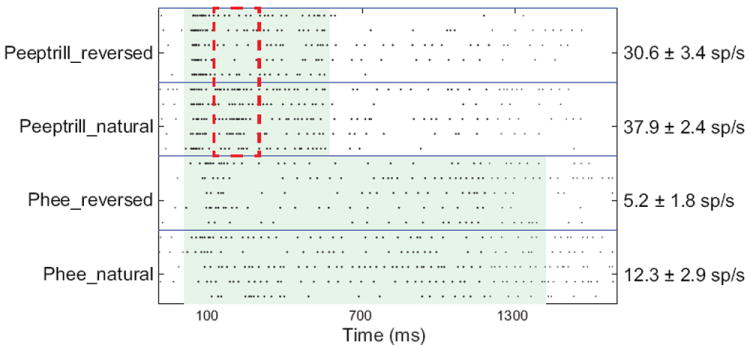
References
-
- Aitkin LM, Dunlop CW, Webster WR. Click-evoked response patterns of single units in the medial geniculate body of the cat. Journal of Neurophysiology. 1966;29:109–123. - PubMed
-
- Aitkin LM, Prain SM. Medial geniculate body: unit responses in the awake cat. Journal of Neurophysiology. 1974;37:512–521. - PubMed
-
- Aitkin L. Rate-level functions of neurons in the inferior colliculus of cats measured with the use of free-field sound stimuli. Journal of Neurophysiology. 1991;65:383–92. - PubMed
-
- Allon N, Yeshurun Y, Wollberg Z. Responses of single cells in the medial geniculate body of awake squirrel monkeys. Experimental Brain Research. 1981;41:222–232. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
