

A cytoprotective perspective on longevity regulation
- PMID: 23726168
- PMCID: PMC4057428
- DOI: 10.1016/j.tcb.2013.04.007
A cytoprotective perspective on longevity regulation
Abstract
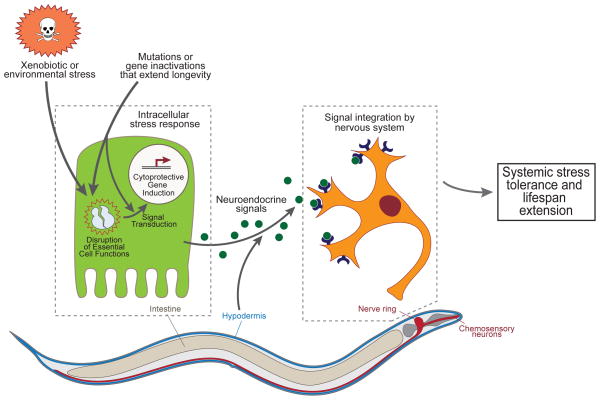
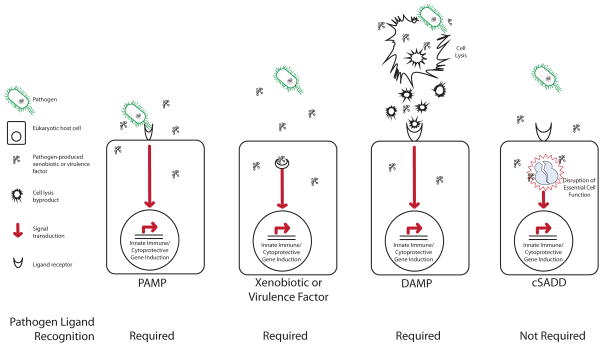
There are many mechanisms of lifespan extension, including the disruption of insulin/insulin-like growth factor 1 (IGF-1) signaling, metabolism, translation, and feeding. Despite the disparate functions of these pathways, inhibition of each induces responses that buffer stress and damage. Here, emphasizing data from genetic analyses in Caenorhabditis elegans, we explore the effectors and upstream regulatory components of numerous cytoprotective mechanisms activated as major elements of longevity programs, including detoxification, innate immunity, proteostasis, and oxidative stress response. We show that their induction underpins longevity extension across functionally diverse triggers and across species. Intertwined with the evolution of longevity, cytoprotective pathways are coupled to the surveillance of core cellular components, with important implications in normal and aberrant responses to drugs, chemicals, and pathogens.
Keywords: aging; cytoprotection; detoxification; hormesis; longevity; stress.
Copyright © 2013. Published by Elsevier Ltd.
Figures
References
-
- Lyman CP, et al. Hibernation and longevity in the Turkish hamster Mesocricetus brandti. Science. 1981;212:668–670. - PubMed
-
- Tatar M, et al. Negligible Senescence during Reproductive Dormancy in Drosophila melanogaster. Am Nat. 2001;158:248–258. - PubMed
-
- Houthoofd K, et al. Ageing is reversed, and metabolism is reset to young levels in recovering dauer larvae of C. elegans. Exp Gerontol. 2002;37:1015–1021. - PubMed
-
- Wilkinson G, South J. Life history, ecology and longevity in bats. Aging Cell. 2002;1:124–155. - PubMed
-
- Cassada RC, Russell RL. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev Biol. 1975;46:326–342. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
