

Nucleosome dynamics regulates DNA processing
- PMID: 23728291
- PMCID: PMC3711194
- DOI: 10.1038/nsmb.2585
Nucleosome dynamics regulates DNA processing
Abstract
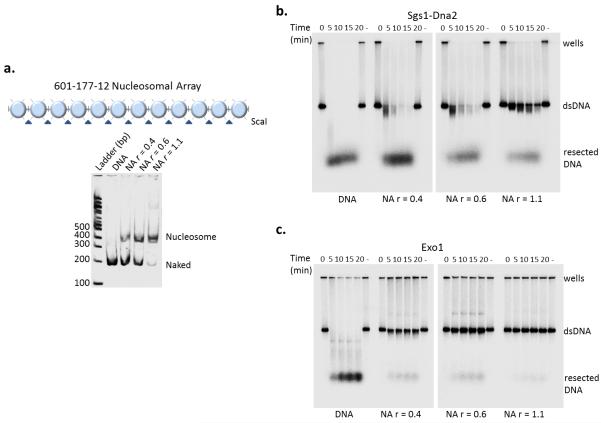
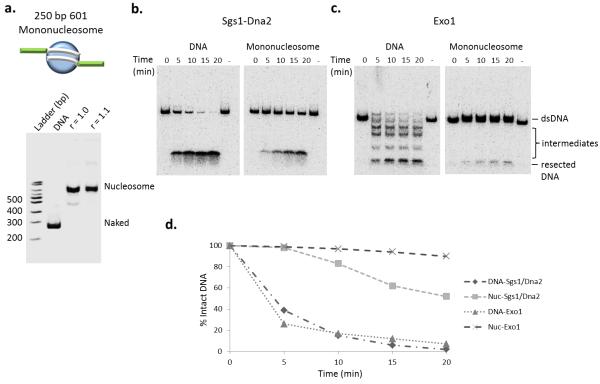
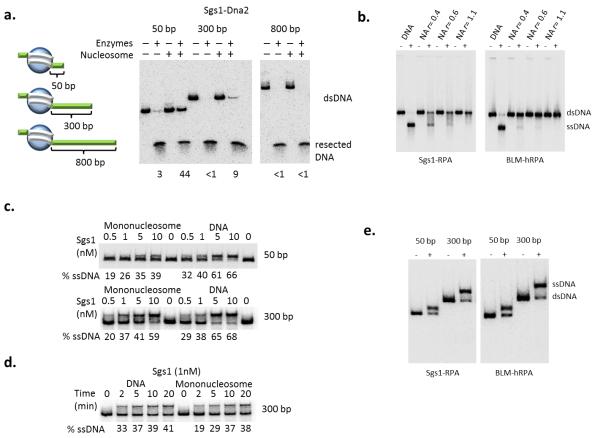
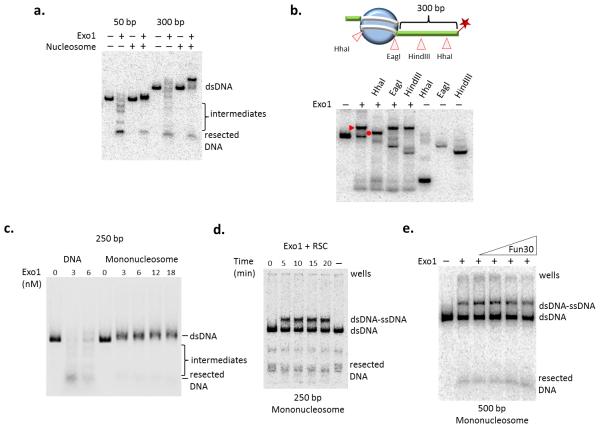
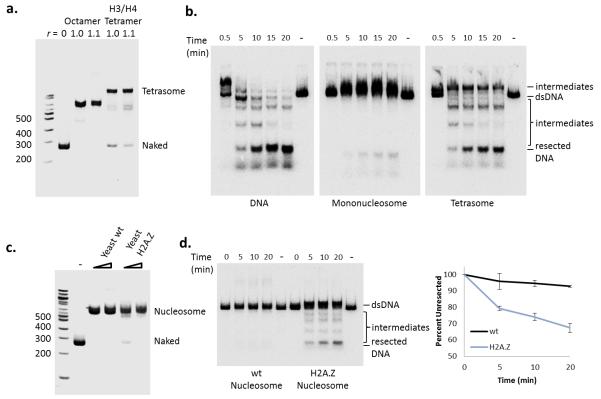
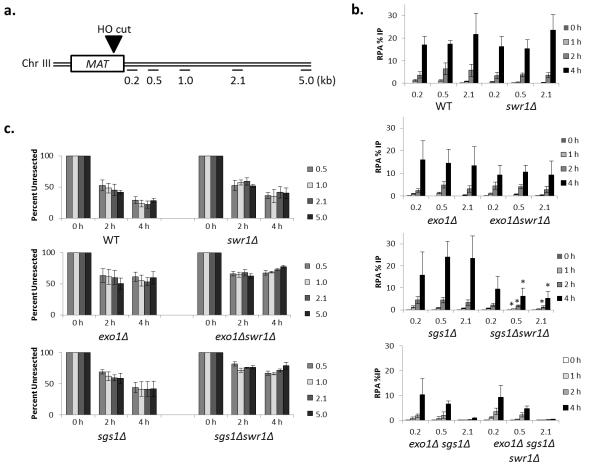
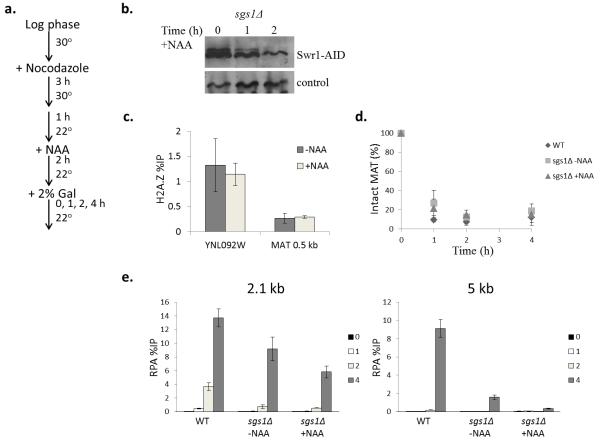
The repair of DNA double-strand breaks (DSBs) is critical for the maintenance of genome integrity. The first step in DSB repair by homologous recombination is the processing of the ends by one of two resection pathways, executed by the Saccharomyces cerevisiae Exo1 and Sgs1-Dna2 machineries. Here we report in vitro and in vivo studies that characterize the impact of chromatin on each resection pathway. We find that efficient resection by the Sgs1-Dna2-dependent machinery requires a nucleosome-free gap adjacent to the DSB. Resection by Exo1 is blocked by nucleosomes, and processing activity can be partially restored by removal of the H2A-H2B dimers. Our study also supports a role for the dynamic incorporation of the H2A.Z histone variant in Exo1 processing, and it further suggests that the two resection pathways require distinct chromatin remodeling events to navigate chromatin structure.
Figures
References
-
- Khanna KK, Jackson SP. DNA double-strand breaks: signaling, repair and the cancer connection. Nat Genet. 2001;27:247–254. - PubMed
-
- Zou L, Elledge SJ. Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes. Science. 2003;300:1542–1548. - PubMed
-
- Krogh BO, Symington LS. Recombination proteins in yeast. Annu Rev Genet. 2004;38:233–271. - PubMed
Methods-only reference
-
- Sigurdsson S, Trujillo K, Song B, Stratton S, Sung P. Basis for avid homologous DNA strand exchange by human Rad51 and RPA. J Biol Chem. 2001;276:8798–8806. - PubMed
-
- Raynard S, Bussen W, Sung PA. A double Holliday junction dissolvasome comprising BLM, topoisomerase III alpha, and BLAP75. J Biol Chem. 2006;281:13861–13864. - PubMed
-
- Luger K, Rechsteiner TJ, Richmond TJ. Preparation of nucleosome core particle from recombinant histones. Methods Enzymol. 1999;304:3–19. - PubMed
-
- Smith CL, Horowitz-Scherer R, Flanagan JF, Woodcock CL, Peterson CL. Structural analysis of the yeast SWI/SNF chromatin remodeling complex. Nat Struct Biol. 2003;10:141–145. - PubMed
-
- Goldstein AL, McCusker JH. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast. 1999;15:1541–1553. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
