

BMI1 represses Ink4a/Arf and Hox genes to regulate stem cells in the rodent incisor
- PMID: 23728424
- PMCID: PMC3735916
- DOI: 10.1038/ncb2766
BMI1 represses Ink4a/Arf and Hox genes to regulate stem cells in the rodent incisor
Abstract
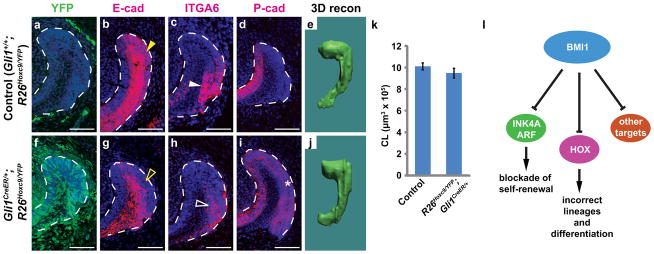
The polycomb group gene Bmi1 is required for maintenance of adult stem cells in many organs. Inactivation of Bmi1 leads to impaired stem cell self-renewal due to deregulated gene expression. One critical target of BMI1 is Ink4a/Arf, which encodes the cell-cycle inhibitors p16(Ink4a) and p19(Arf). However, deletion of Ink4a/Arf only partially rescues Bmi1-null phenotypes, indicating that other important targets of BMI1 exist. Here, using the continuously growing mouse incisor as a model system, we report that Bmi1 is expressed by incisor stem cells and that deletion of Bmi1 resulted in fewer stem cells, perturbed gene expression and defective enamel production. Transcriptional profiling revealed that Hox expression is normally repressed by BMI1 in the adult, and functional assays demonstrated that BMI1-mediated repression of Hox genes preserves the undifferentiated state of stem cells. As Hox gene upregulation has also been reported in other systems when Bmi1 is inactivated, our findings point to a general mechanism whereby BMI1-mediated repression of Hox genes is required for the maintenance of adult stem cells and for prevention of inappropriate differentiation.
Figures
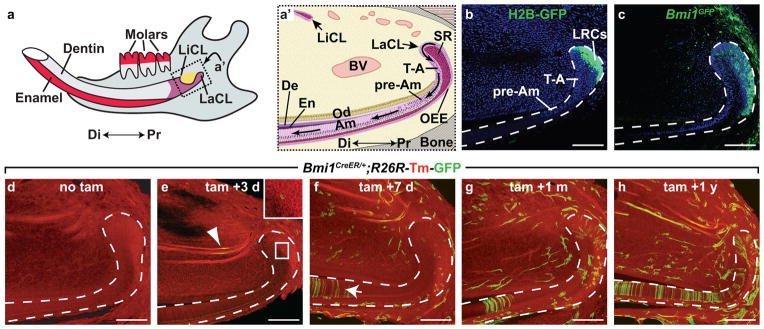
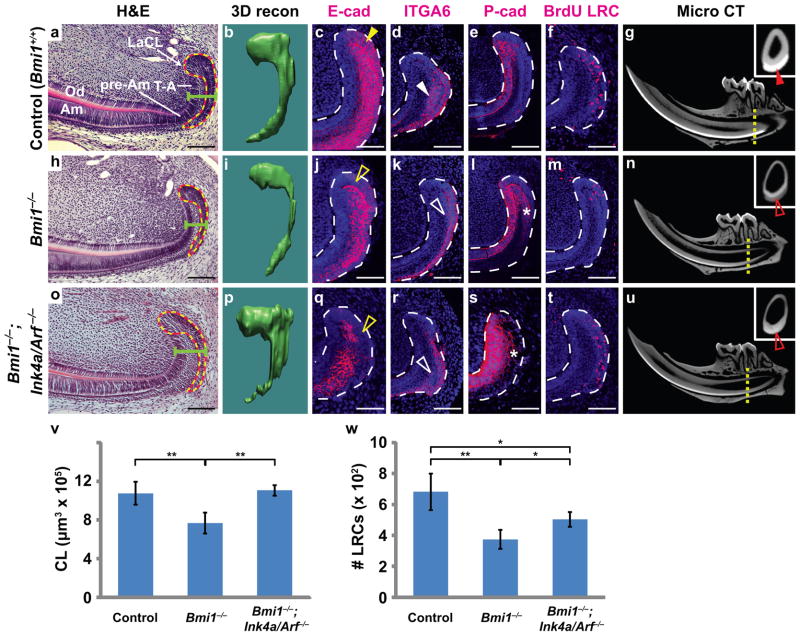
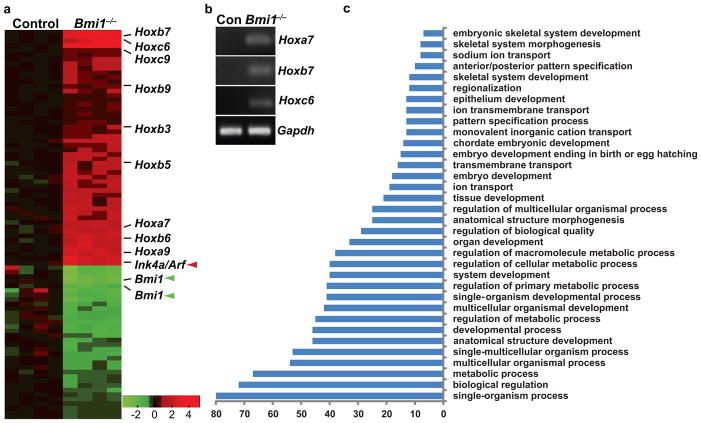
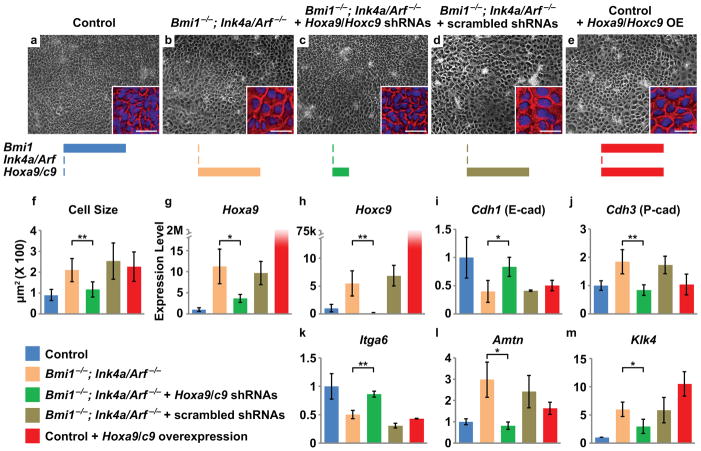
References
-
- Park IK, et al. Bmi-1 is required for maintenance of adult self-renewing haematopoietic stem cells. Nature. 2003;423:302–305. - PubMed
-
- Jacobs JJ, Kieboom K, Marino S, DePinho RA, van Lohuizen M. The oncogene and Polycomb-group gene bmi-1 regulates cell proliferation and senescence through the ink4a locus. Nature. 1999;397:164–168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
