

Ezrin/radixin/moesin proteins differentially regulate endothelial hyperpermeability after thrombin
- PMID: 23729486
- PMCID: PMC3743011
- DOI: 10.1152/ajplung.00355.2012
Ezrin/radixin/moesin proteins differentially regulate endothelial hyperpermeability after thrombin
Abstract
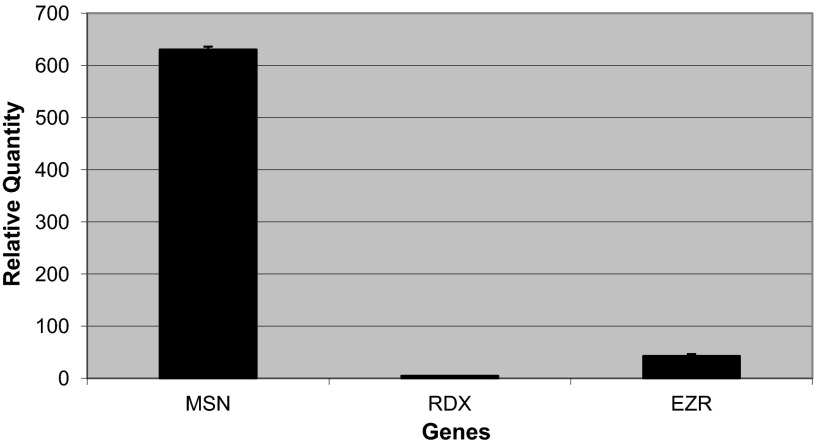
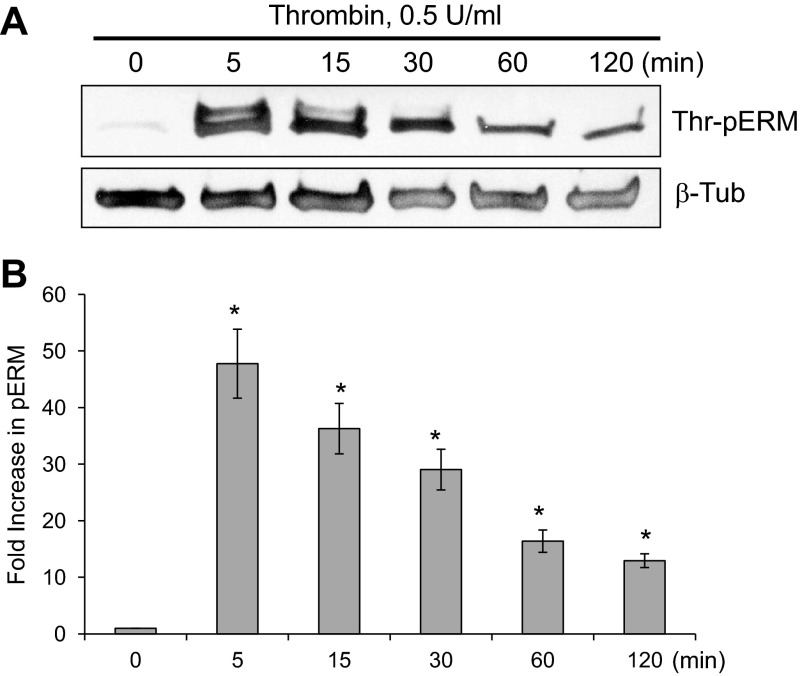
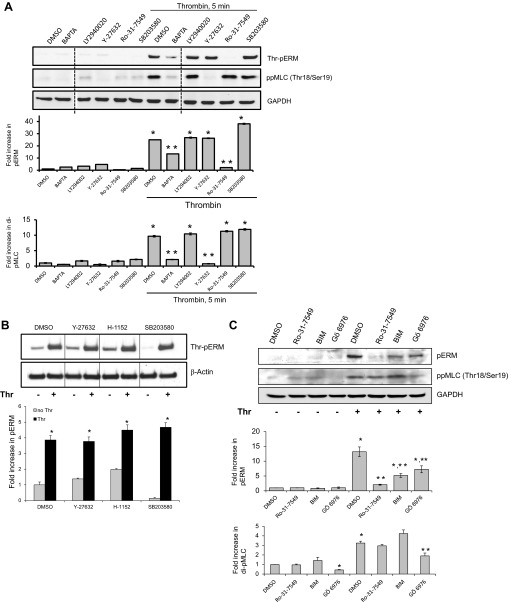
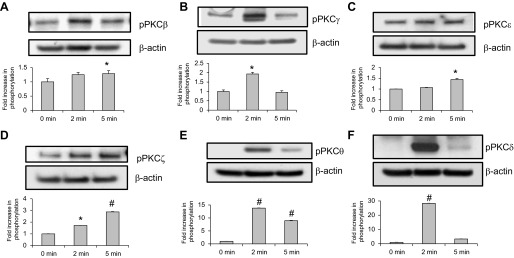
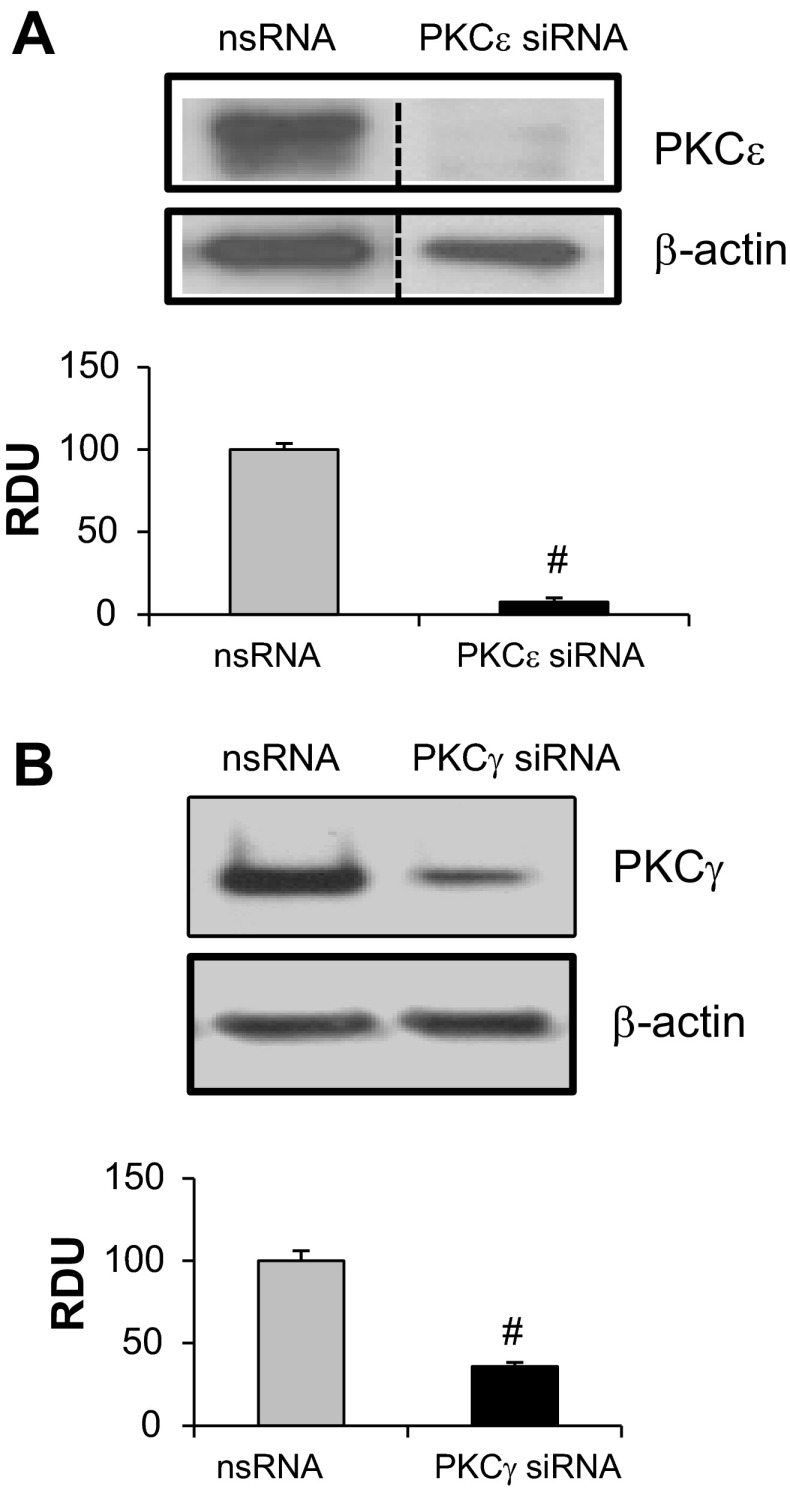
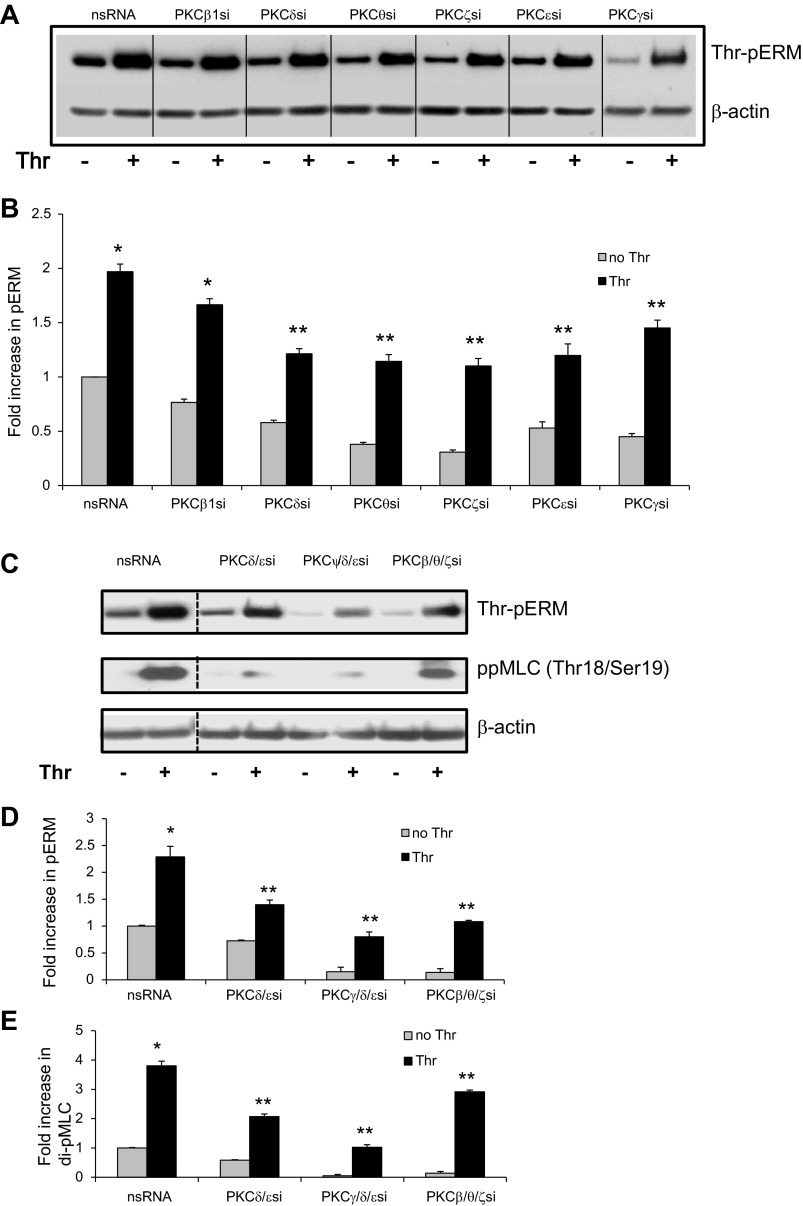
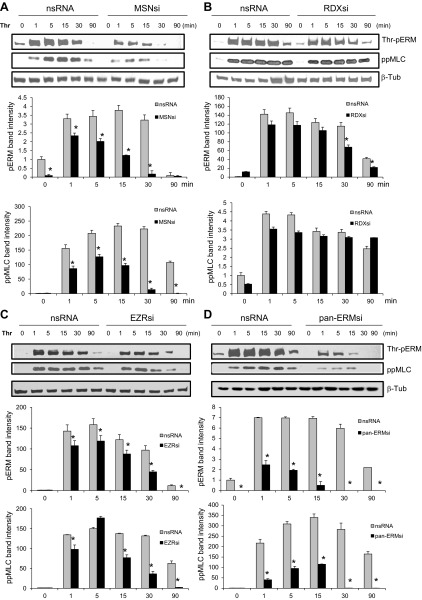
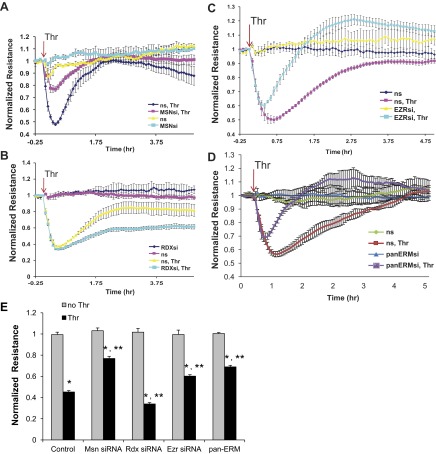
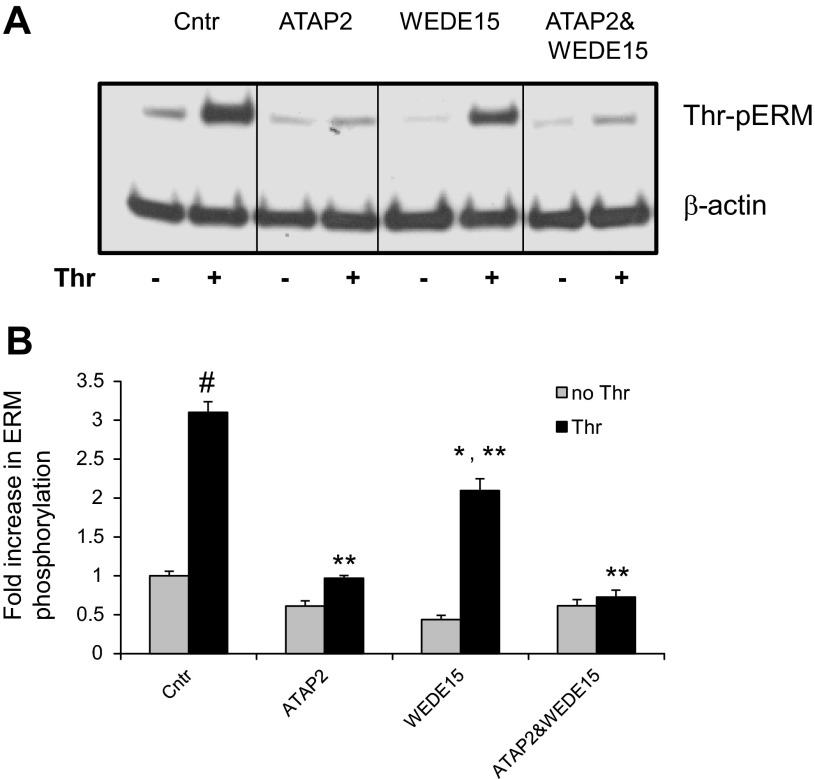
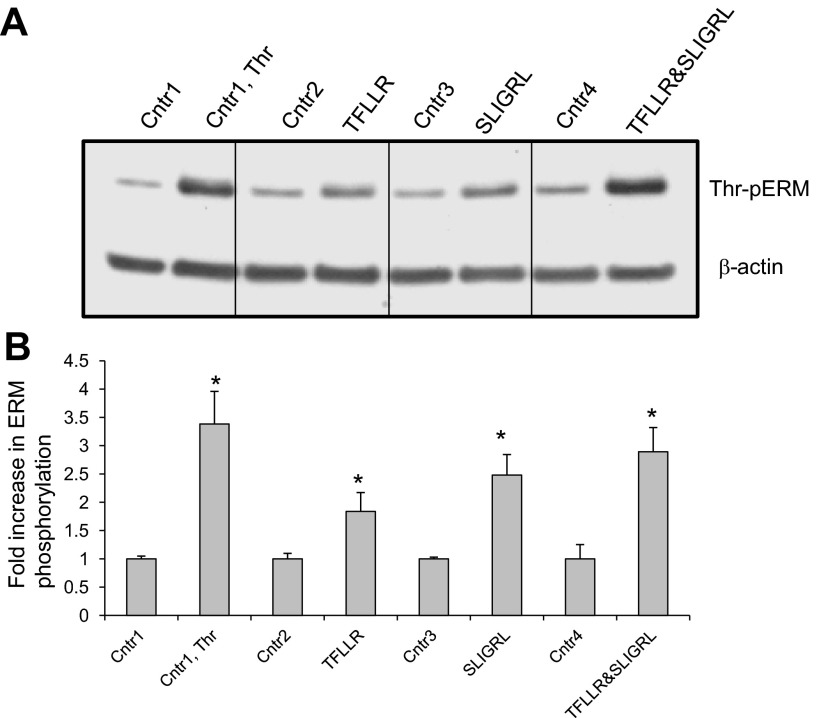
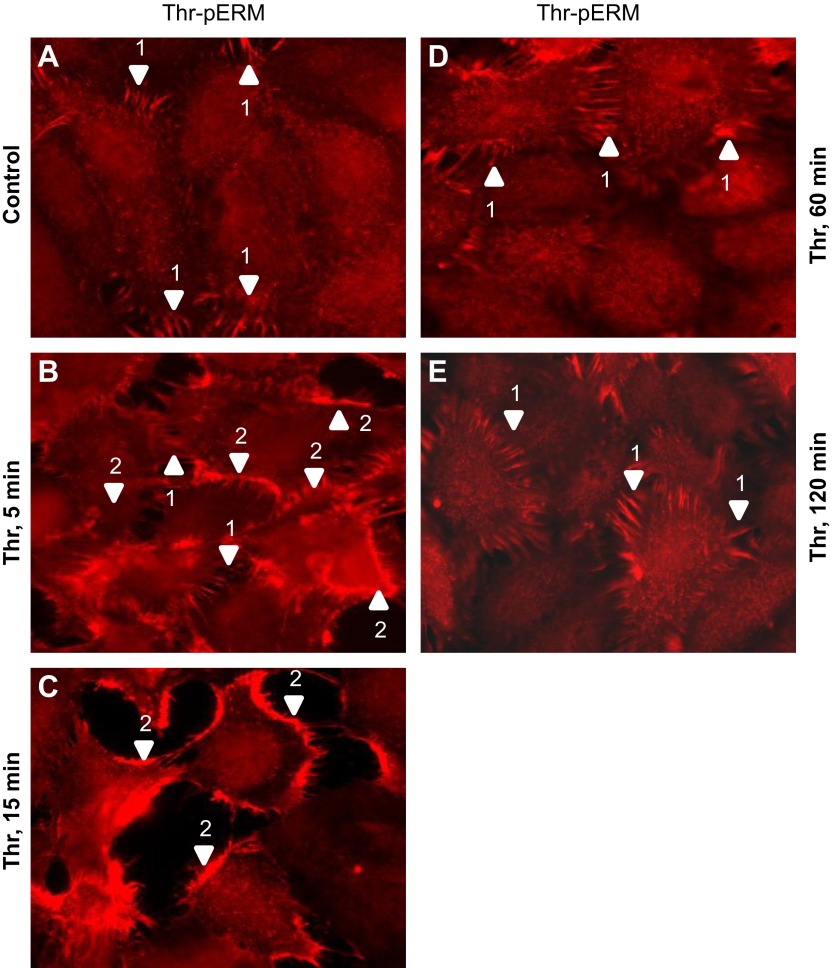
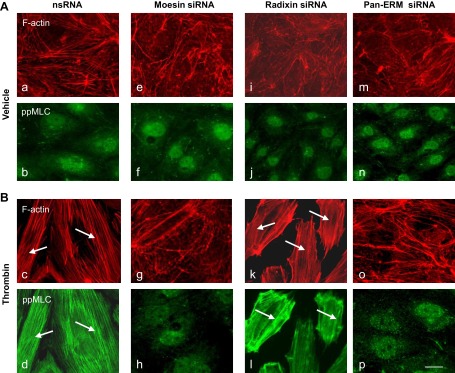
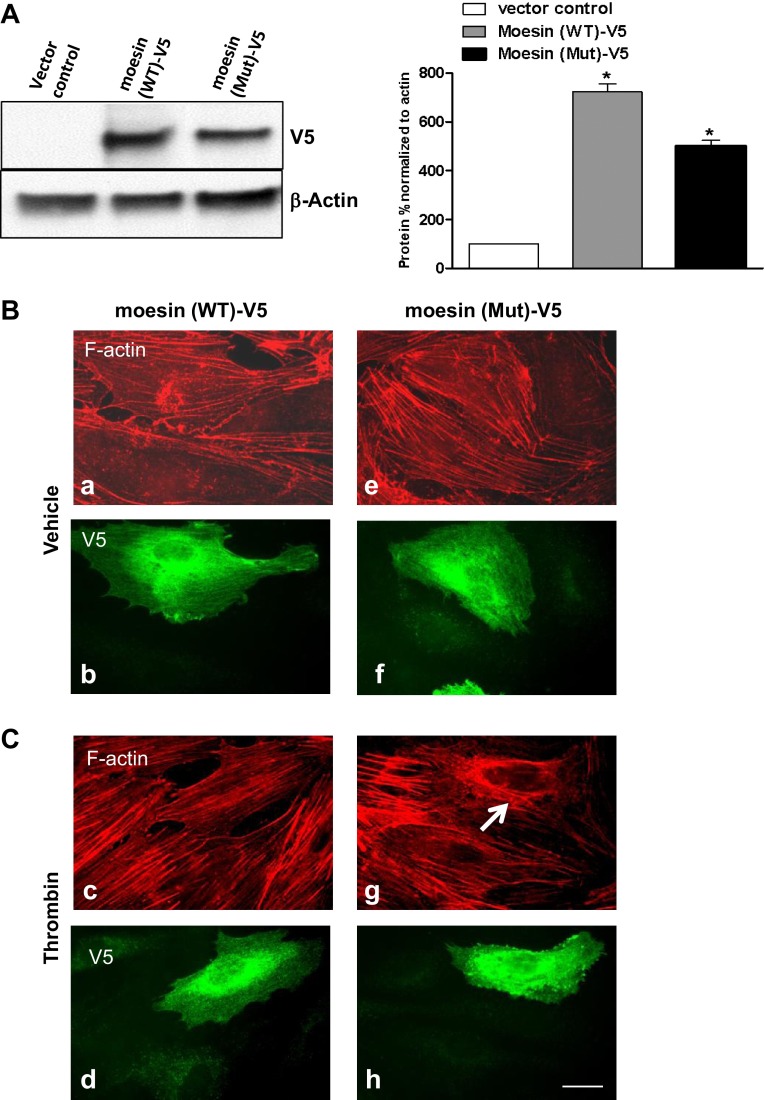
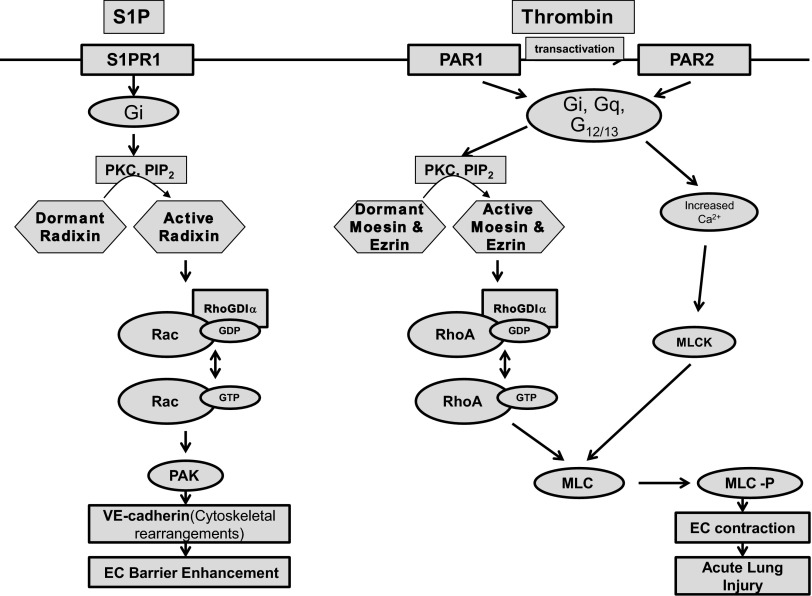
Endothelial cell (EC) barrier disruption induced by inflammatory agonists such as thrombin leads to potentially lethal physiological dysfunction such as alveolar flooding, hypoxemia, and pulmonary edema. Thrombin stimulates paracellular gap and F-actin stress fiber formation, triggers actomyosin contraction, and alters EC permeability through multiple mechanisms that include protein kinase C (PKC) activation. We previously have shown that the ezrin, radixin, and moesin (ERM) actin-binding proteins differentially participate in sphingosine-1 phosphate-induced EC barrier enhancement. Phosphorylation of a conserved threonine residue in the COOH-terminus of ERM proteins causes conformational changes in ERM to unmask binding sites and is considered a hallmark of ERM activation. In the present study we test the hypothesis that ERM proteins are phosphorylated on this critical threonine residue by thrombin-induced signaling events and explore the role of the ERM family in modulating thrombin-induced cytoskeletal rearrangement and EC barrier function. Thrombin promotes ERM phosphorylation at this threonine residue (ezrin Thr567, radixin Thr564, moesin Thr558) in a PKC-dependent fashion and induces translocation of phosphorylated ERM to the EC periphery. Thrombin-induced ERM threonine phosphorylation is likely synergistically mediated by protease-activated receptors PAR1 and PAR2. Using the siRNA approach, depletion of either moesin alone or of all three ERM proteins significantly attenuates thrombin-induced increase in EC barrier permeability (transendothelial electrical resistance), cytoskeletal rearrangements, paracellular gap formation, and accumulation of phospho-myosin light chain. In contrast, radixin depletion exerts opposing effects on these indexes. These data suggest that ERM proteins play important differential roles in the thrombin-induced modulation of EC permeability, with moesin promoting barrier dysfunction and radixin opposing it.
Keywords: ERM; PKC; barrier dysfunction; cytoskeleton; endothelial cells; phosphorylation; thrombin.
Figures
References
-
- Berryman M, Franck Z, Bretscher A. Ezrin is concentrated in the apical microvilli of a wide variety of epithelial cells whereas moesin is found primarily in endothelial cells. J Cell Sci 105: 1025–1043, 1993 - PubMed
-
- Birukova AA, Birukov KG, Smurova K, Adyshev D, Kaibuchi K, Alieva I, Garcia JG, Verin AD. Novel role of microtubules in thrombin-induced endothelial barrier dysfunction. FASEB J 18: 1879–1890, 2004 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous