

SA1 binds directly to DNA through its unique AT-hook to promote sister chromatid cohesion at telomeres
- PMID: 23729739
- PMCID: PMC3730250
- DOI: 10.1242/jcs.130872
SA1 binds directly to DNA through its unique AT-hook to promote sister chromatid cohesion at telomeres
Abstract
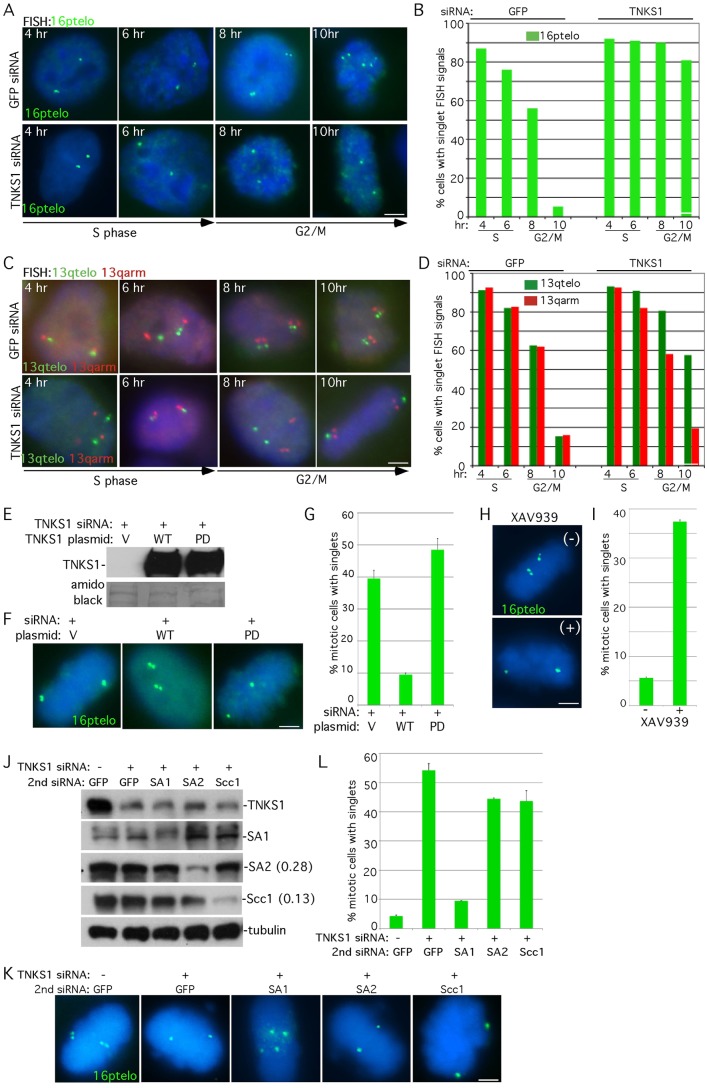
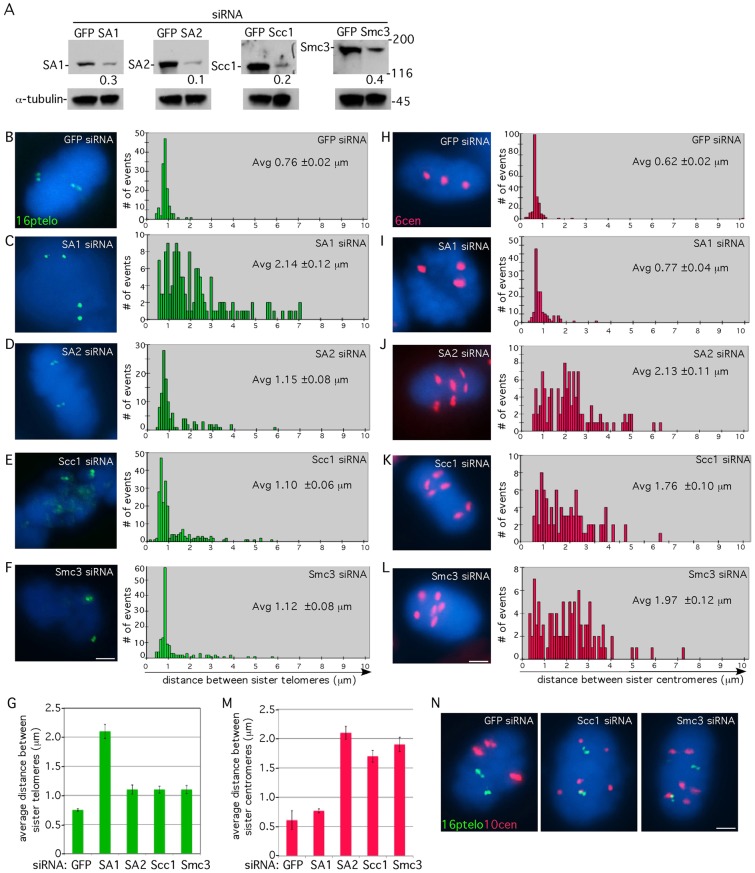
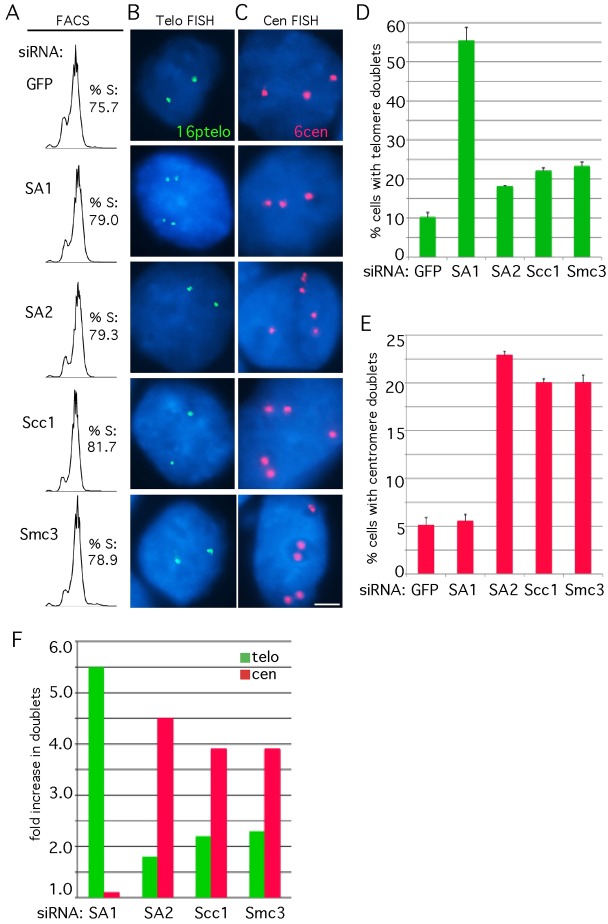
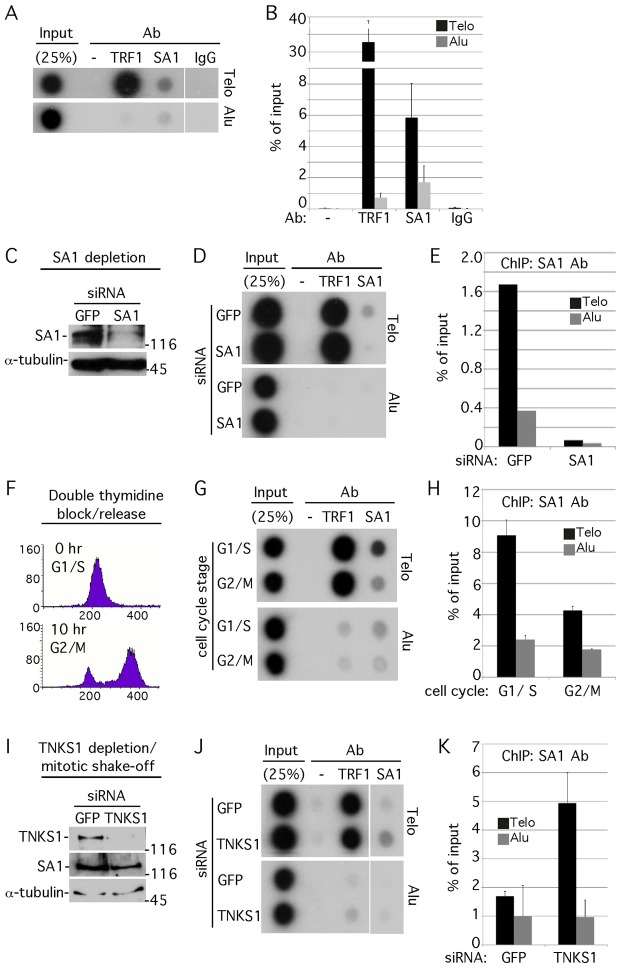
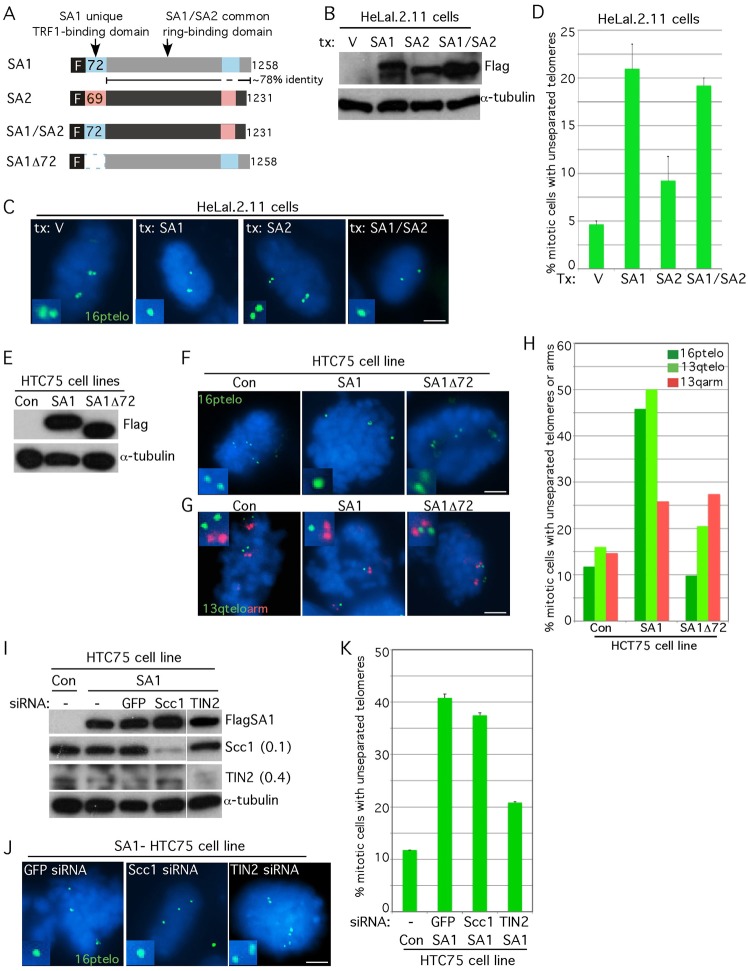
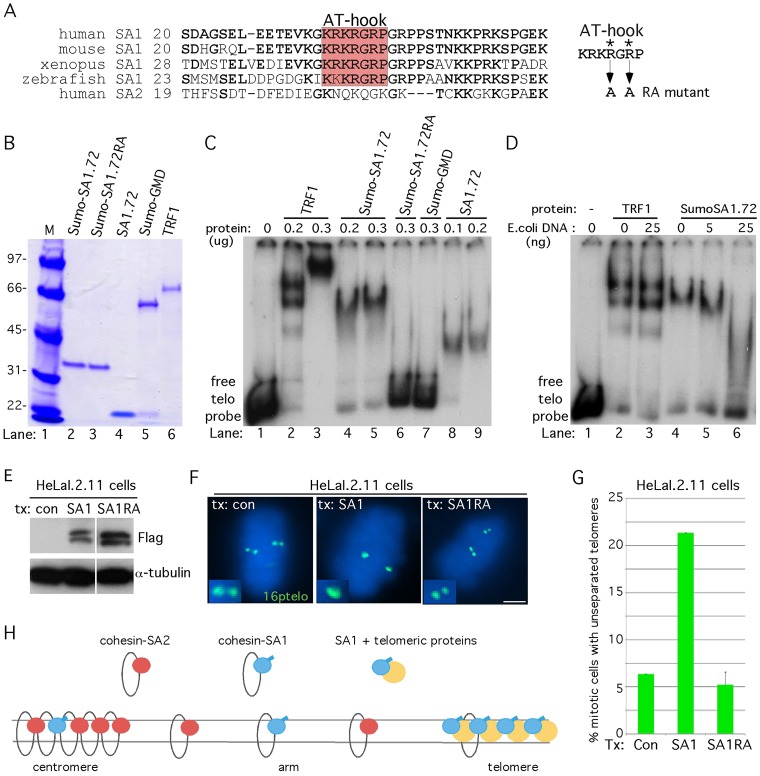
Sister chromatid cohesion relies on cohesin, a complex comprising a tri-partite ring and a peripheral subunit Scc3, which is found as two related isoforms SA1 and SA2 in vertebrates. There is a division of labor between the vertebrate cohesin complexes; SA1-cohesin is required at telomeres and SA2-cohesin at centromeres. Depletion of SA1 has dramatic consequences for telomere function and genome integrity, but the mechanism by which SA1-cohesin mediates cohesion at telomeres is not well understood. Here we dissect the individual contribution of SA1 and the ring subunits to telomere cohesion and show that telomeres rely heavily on SA1 and to a lesser extent on the ring for cohesion. Using chromatin immunoprecipitation we show that SA1 is highly enriched at telomeres, is decreased at mitosis when cohesion is resolved, and is increased when cohesion persists. Overexpression of SA1 alone was sufficient to induce cohesion at telomeres, independent of the cohesin ring and dependent on its unique (not found in SA2) N-terminal domain, which we show binds to telomeric DNA through an AT-hook motif. We suggest that a specialized cohesion mechanism may be required to accommodate the high level of DNA replication-associated repair at telomeres.
Keywords: Cohesion; SA1; Telomeres.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
