

Gallbladder epithelium as a niche for chronic Salmonella carriage
- PMID: 23732169
- PMCID: PMC3719562
- DOI: 10.1128/IAI.00258-13
Gallbladder epithelium as a niche for chronic Salmonella carriage
Abstract
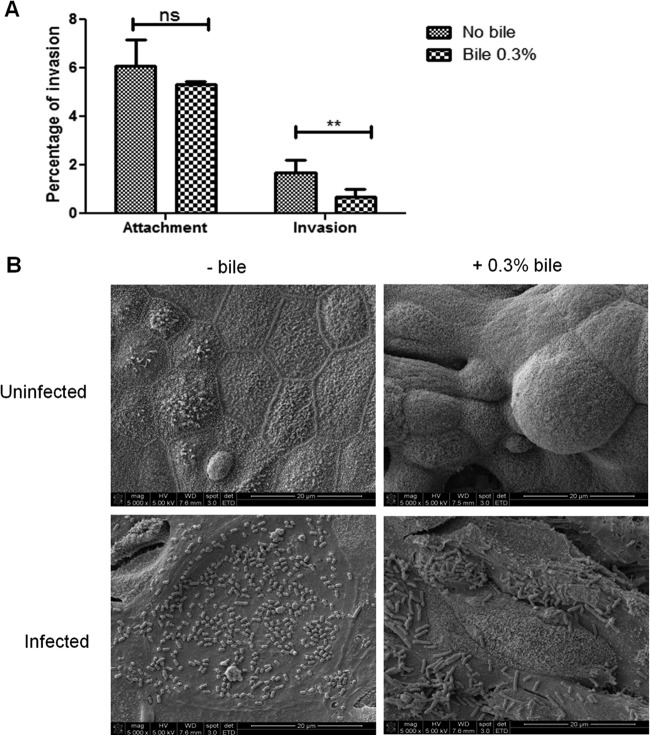
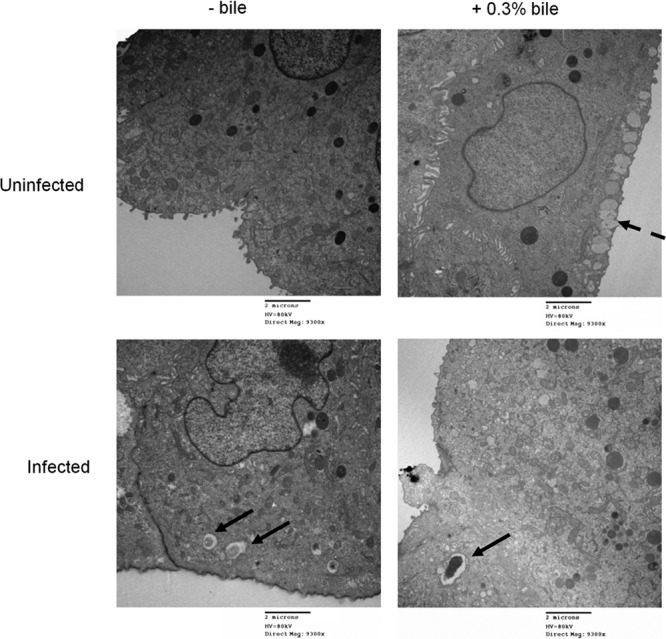
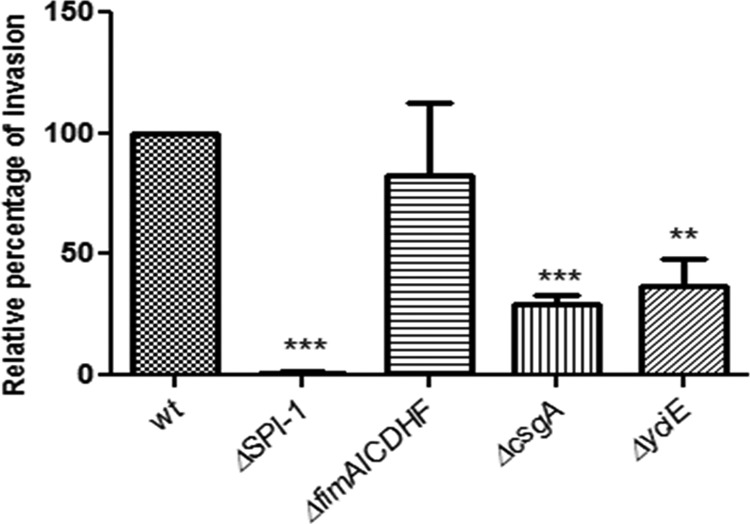
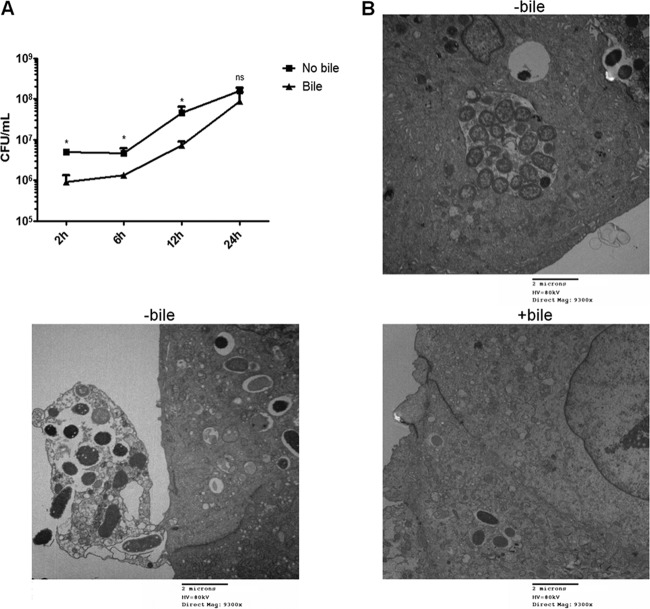
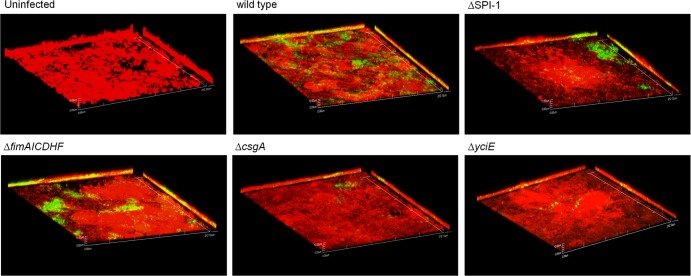
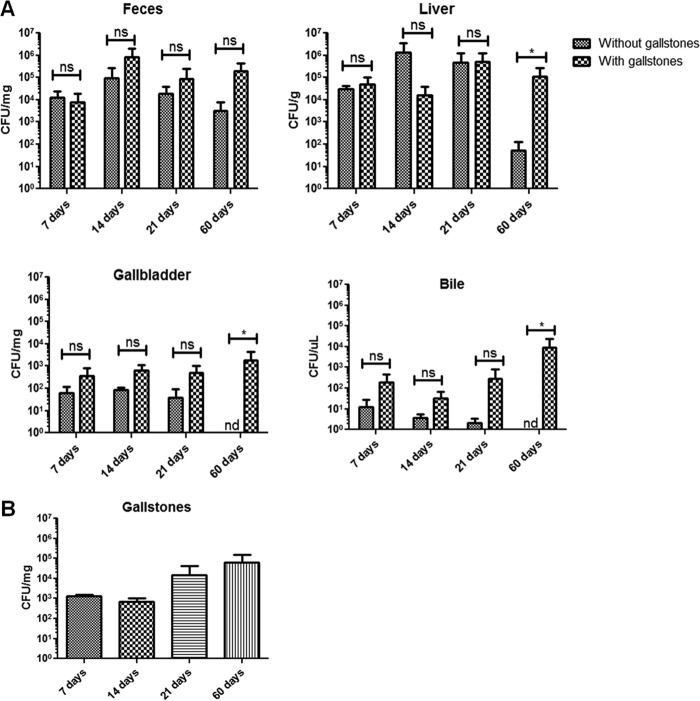
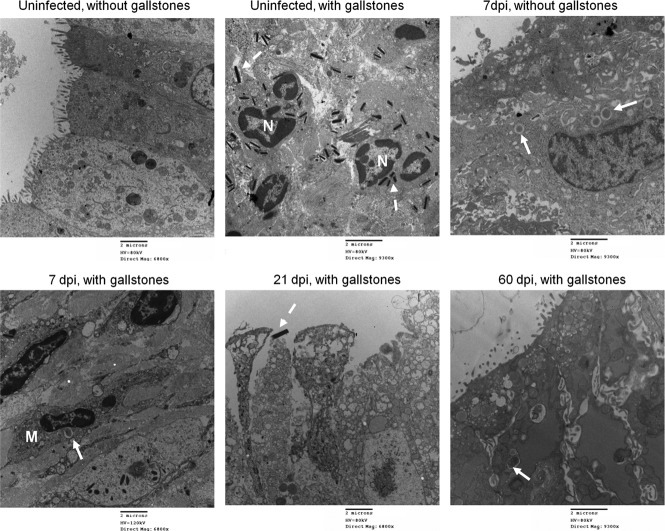
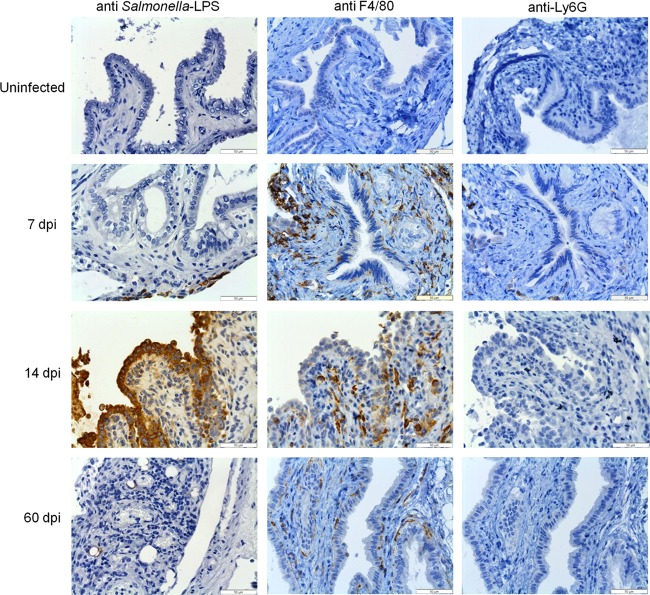
Although typhoid fever has been intensively studied, chronic typhoid carriage still represents a problem for the transmission and persistence of the disease in areas of endemicity. This chronic state is highly associated with the presence of gallstones in the gallbladder of infected carriers upon which Salmonella can form robust biofilms. However, we hypothesize that in addition to gallstones, the gallbladder epithelium aids in the establishment/maintenance of chronic carriage. In this work, we present evidence of the role of the gallbladder epithelium in chronic carriage by a mechanism involving invasion, intracellular persistence, and biofilm formation. Salmonella was able to adhere to and invade polarized gallbladder epithelial cells apically in the absence and presence of bile in a Salmonella pathogenicity island 1 (SPI-1)-dependent manner. Intracellular replication of Salmonella was also evident at 12 and 24 h postinvasion. A flowthrough system revealed that Salmonella is able to adhere to and form extensive bacterial foci on gallbladder epithelial cells as early as 12 h postinoculation. In vivo experiments using a chronic mouse model of typhoid carriage showed invasion and damage of the gallbladder epithelium and lamina propria up to 2 months after Salmonella infection, with an abundant presence of macrophages, a relative absence of neutrophils, and extrusion of infected epithelial cells. Additionally, microcolonies of Salmonella cells were evident on the surface of the mouse gallbladder epithelia up to 21 days postinfection. These data reveal a second potential mechanism, intracellular persistence and/or bacterial aggregation in/on the gallbladder epithelium with luminal cell extrusion, for Salmonella maintenance in the gallbladder.
Figures


References
-
- Vazquez-Torres A, Jones-Carson J, Baumler AJ, Falkow S, Valdivia R, Brown W, Le M, Berggren R, Parks WT, Fang FC. 1999. Extraintestinal dissemination of Salmonella by CD18-expressing phagocytes. Nature 401:804–808 - PubMed
-
- Levine MM, Black RE, Lanata C. 1982. Precise estimation of the numbers of chronic carriers of Salmonella Typhi in Santiago, Chile, an endemic area. J. Infect. Dis. 146:724–726 - PubMed
-
- Merselis JG, Jr, Kaye D, Connolly CS, Hook EW. 1964. Quantitative bacteriology of the typhoid carrier state. Am. J. Trop. Med. Hyg. 13:425–429 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
