

Restricted perinatal retinal degeneration induces retina reshaping and correlated structural rearrangement of the retinotopic map
- PMID: 23733098
- PMCID: PMC3709497
- DOI: 10.1038/ncomms2926
Restricted perinatal retinal degeneration induces retina reshaping and correlated structural rearrangement of the retinotopic map
Abstract
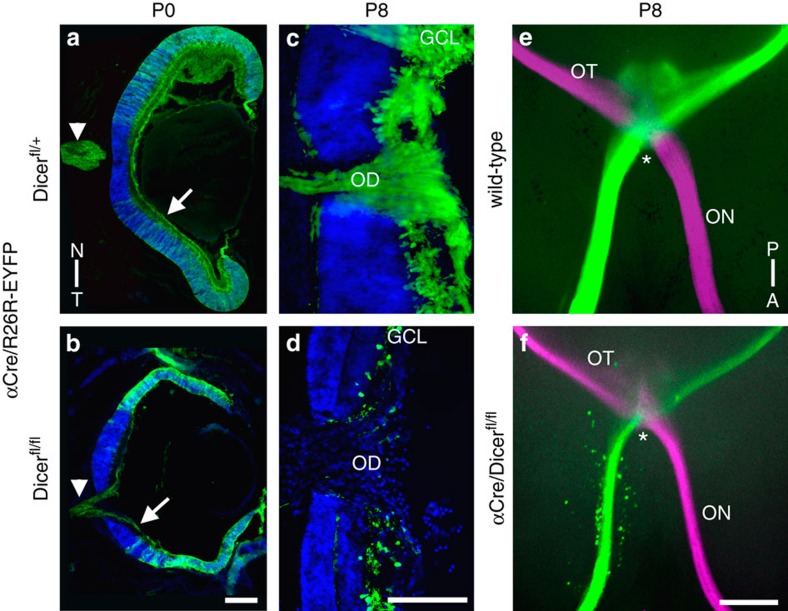
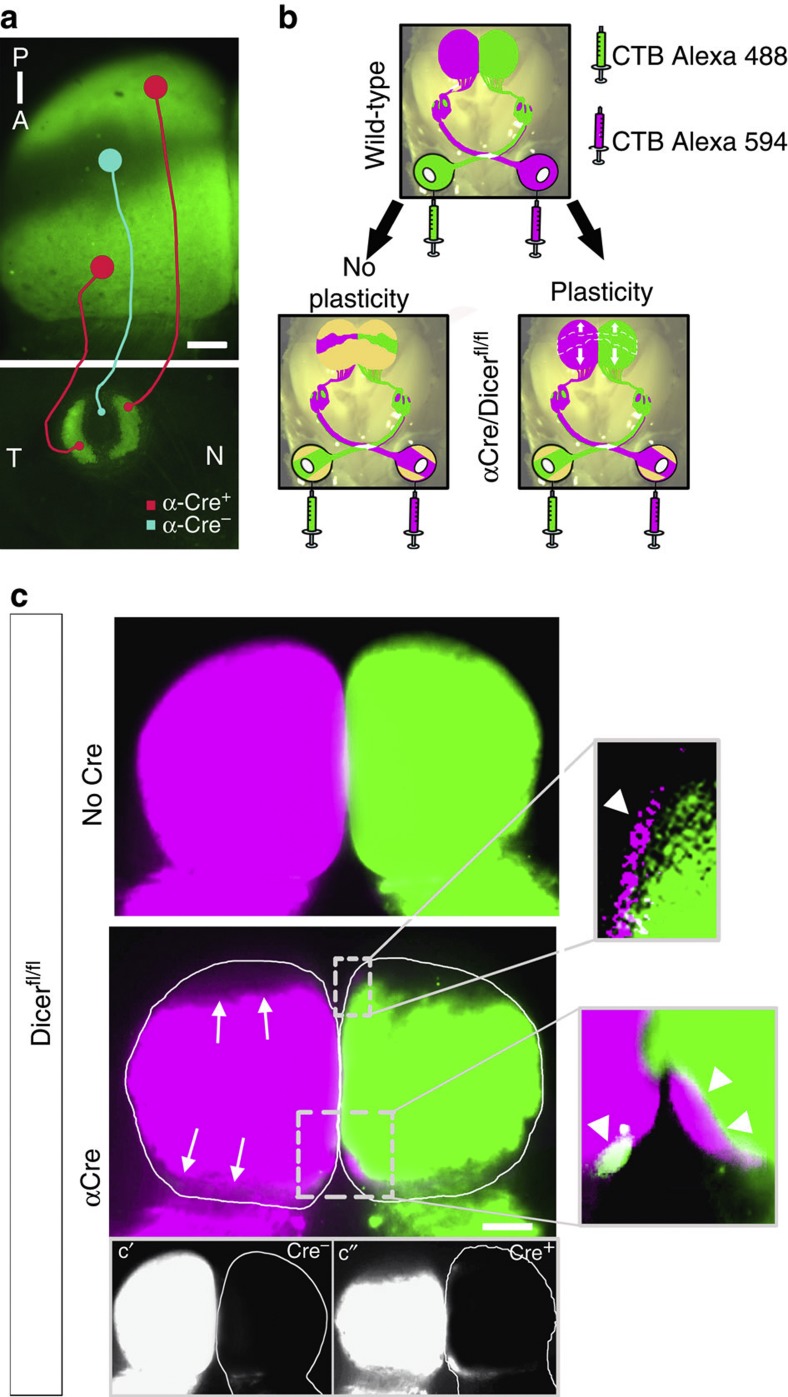
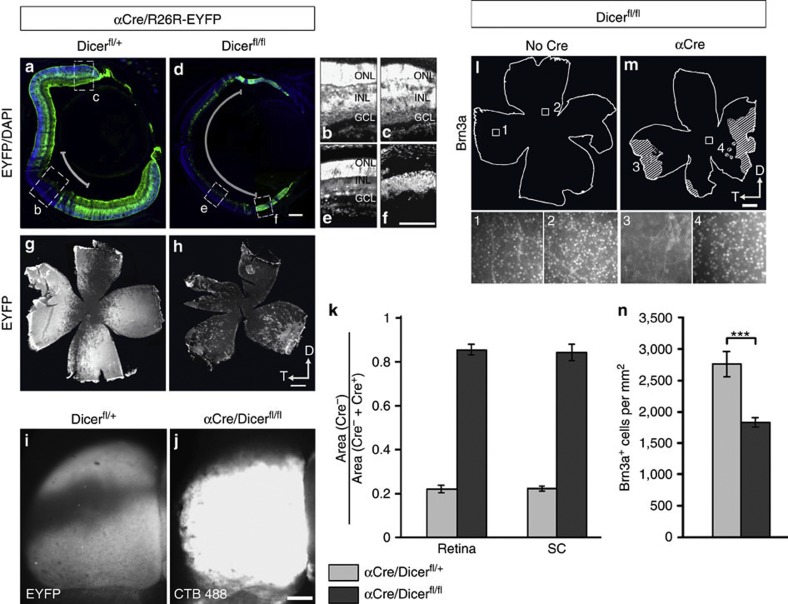
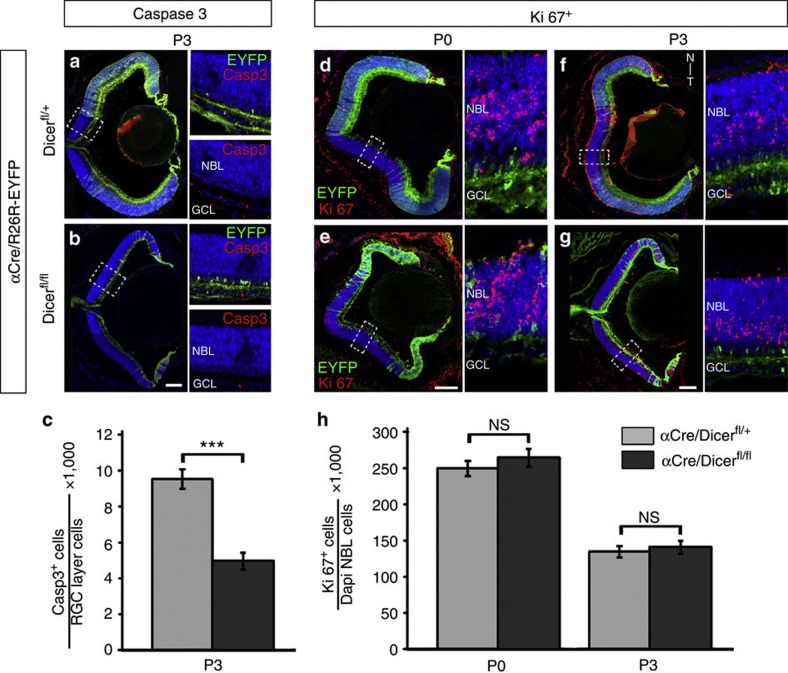
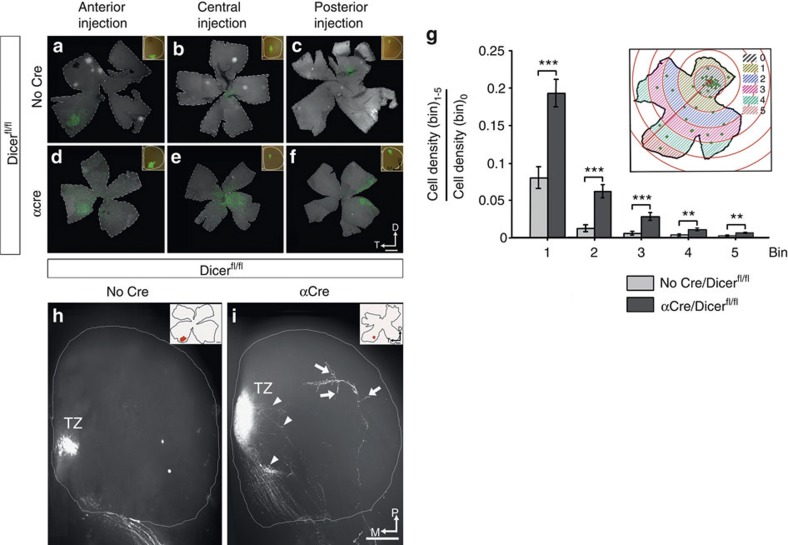
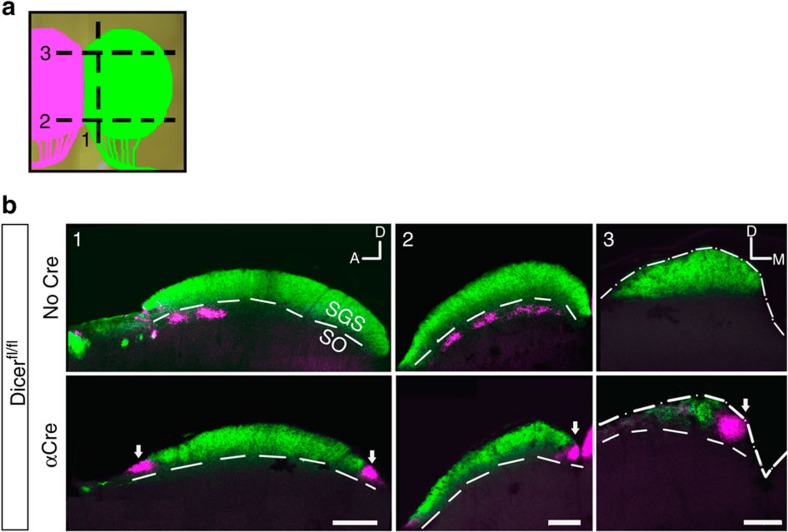
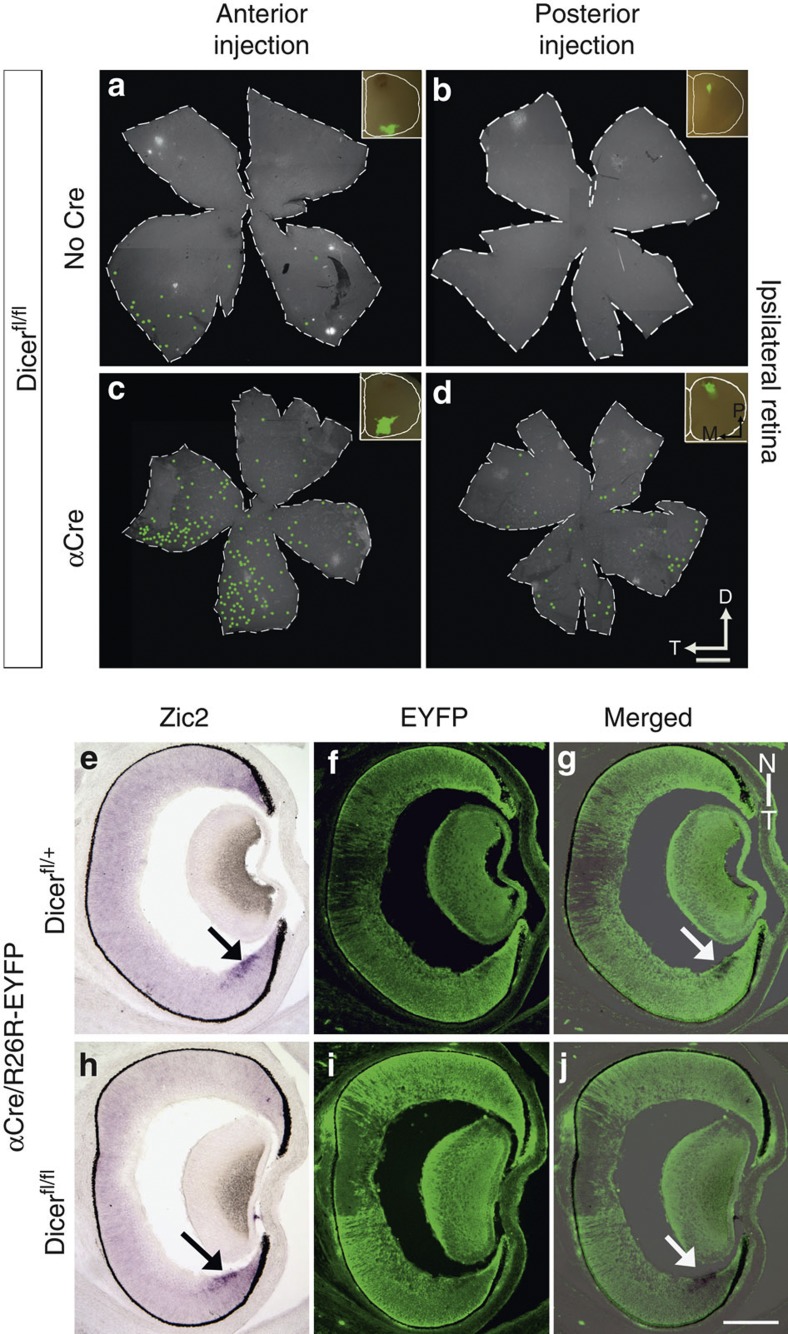
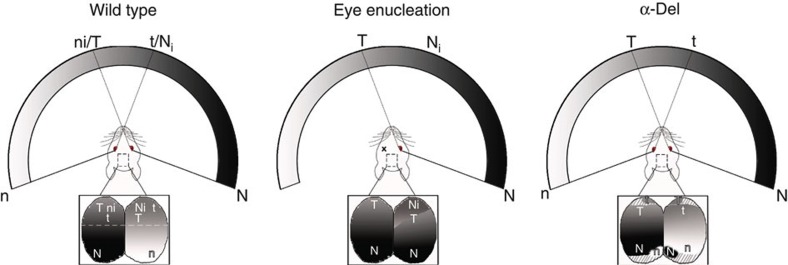
The formation of the retinotopic map depends on the action of axon guidance molecules, activity-dependent mechanisms and axonal competition. However, little is known about the plasticity potential of the system and the effects on the remodelling of retinocollicular connections upon retinal insults. Here we create a mouse model in which retinal ganglion cells that project to anterior and posterior superior colliculus undergo cell death during topographic map formation. We show that the remaining retinal ganglion cells expand the targeted area in the superior colliculus and at the same time increase their spatial coverage in the retina in a correlated fashion. The resulting contralateral topographic map is overall maintained but less precise, while ipsilateral retinal ganglion cell axons are abnormally distributed in anterior and posterior superficial superior colliculus. These results suggest the presence of plastic mechanisms in the developing mammalian visual system to adjust retinal space and its target coverage and ensure a uniform map.
Figures
References
-
- Hindges R., McLaughlin T., Genoud N., Henkemeyer M. & O’Leary D. D.. EphB forward signaling controls directional branch extension and arborization required for dorsal-ventral retinotopic mapping. Neuron 35, 475–487 (2002) . - PubMed
-
- Drager U. C. & Olsen J. F.. Origins of crossed and uncrossed retinal projections in pigmented and albino mice. J. Comp. Neurol. 191, 383–412 (1980) . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
