

Acute myeloid leukemia creates an arginase-dependent immunosuppressive microenvironment
- PMID: 23733335
- PMCID: PMC3731930
- DOI: 10.1182/blood-2013-01-480129
Acute myeloid leukemia creates an arginase-dependent immunosuppressive microenvironment
Abstract
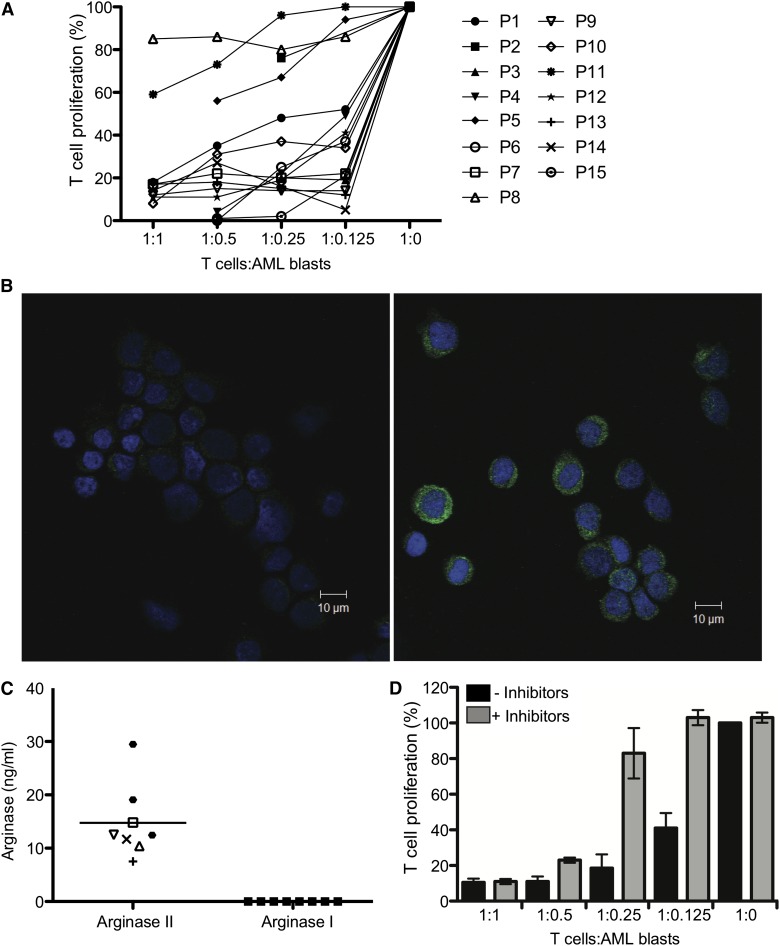
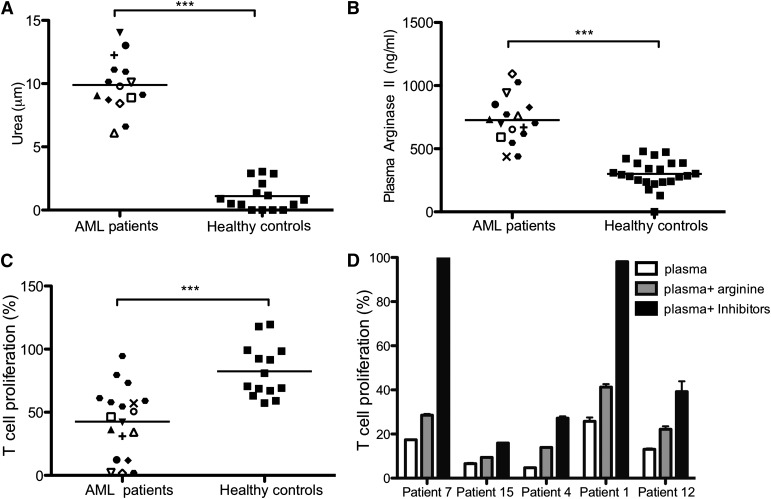
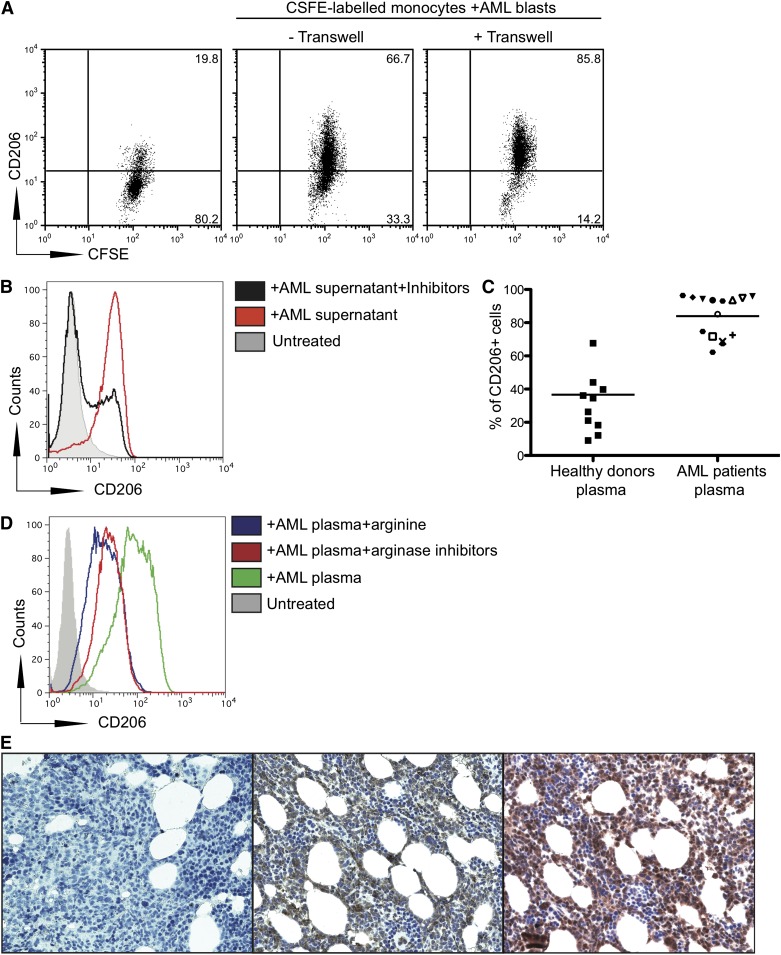
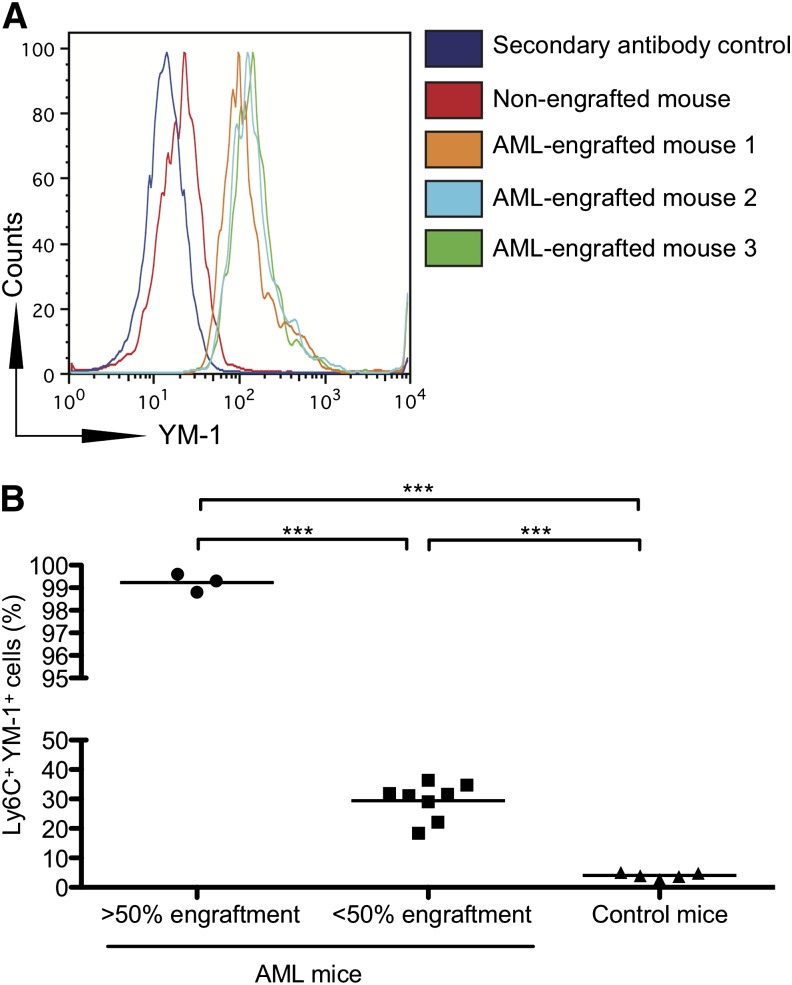
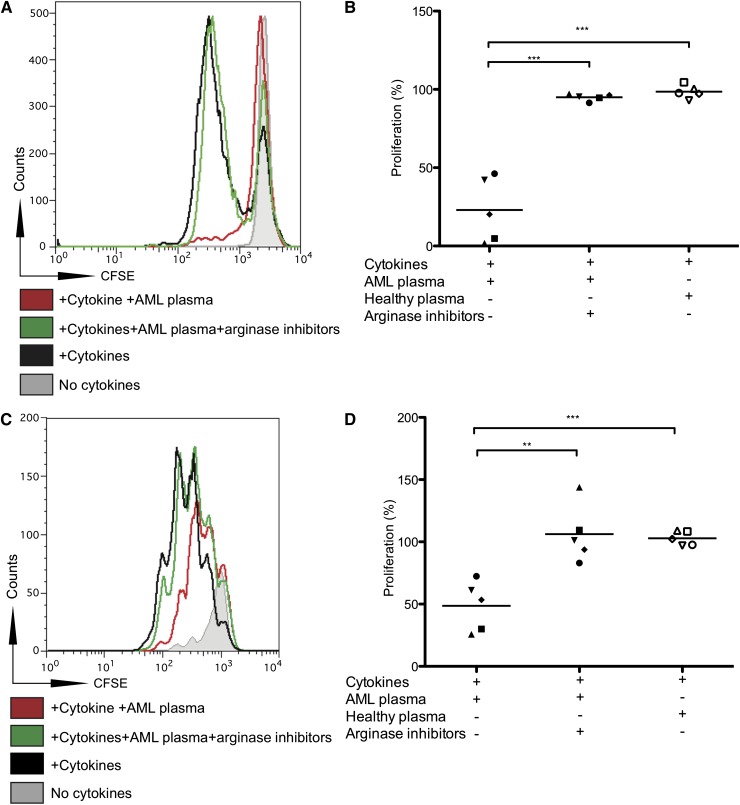
Acute myeloid leukemia (AML) is the most common acute leukemia in adults and the second most common frequent leukemia of childhood. Patients may present with lymphopenia or pancytopenia at diagnosis. We investigated the mechanisms by which AML causes pancytopenia and suppresses patients' immune response. This study identified for the first time that AML blasts alter the immune microenvironment through enhanced arginine metabolism. Arginase II is expressed and released from AML blasts and is present at high concentrations in the plasma of patients with AML, resulting in suppression of T-cell proliferation. We extended these results by demonstrating an arginase-dependent ability of AML blasts to polarize surrounding monocytes into a suppressive M2-like phenotype in vitro and in engrafted nonobese diabetic-severe combined immunodeficiency mice. In addition, AML blasts can suppress the proliferation and differentiation of murine granulocyte-monocyte progenitors and human CD34(+) progenitors. Finally, the study showed that the immunosuppressive activity of AML blasts can be modulated through small-molecule inhibitors of arginase and inducible nitric oxide synthase, suggesting a novel therapeutic target in AML. The results strongly support the hypothesis that AML creates an immunosuppressive microenvironment that contributes to the pancytopenia observed at diagnosis.
Figures
Comment in
-
Arginase-mediated "field" defects in AML.Blood. 2013 Aug 1;122(5):620-1. doi: 10.1182/blood-2013-06-508499. Blood. 2013. PMID: 23908439
References
-
- Marcucci G, Haferlach T, Döhner H. Molecular genetics of adult acute myeloid leukemia: prognostic and therapeutic implications. J Clin Oncol. 2011;29(5):475–486. - PubMed
-
- Woods WG, Neudorf S, Gold S, et al. Children’s Cancer Group. A comparison of allogeneic bone marrow transplantation, autologous bone marrow transplantation, and aggressive chemotherapy in children with acute myeloid leukemia in remission. Blood. 2001;97(1):56–62. - PubMed
-
- Horowitz MM, Gale RP, Sondel PM, et al. Graft-versus-leukemia reactions after bone marrow transplantation. Blood. 1990;75(3):555–562. - PubMed
-
- Gibson BE, Wheatley K, Hann IM, et al. Treatment strategy and long-term results in paediatric patients treated in consecutive UK AML trials. Leukemia. 2005;19(12):2130–2138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
