

Multiple genetic pathways involving amino acid position 143 of HIV-1 integrase are preferentially associated with specific secondary amino acid substitutions and confer resistance to raltegravir and cross-resistance to elvitegravir
- PMID: 23733474
- PMCID: PMC3754334
- DOI: 10.1128/AAC.00204-13
Multiple genetic pathways involving amino acid position 143 of HIV-1 integrase are preferentially associated with specific secondary amino acid substitutions and confer resistance to raltegravir and cross-resistance to elvitegravir
Abstract
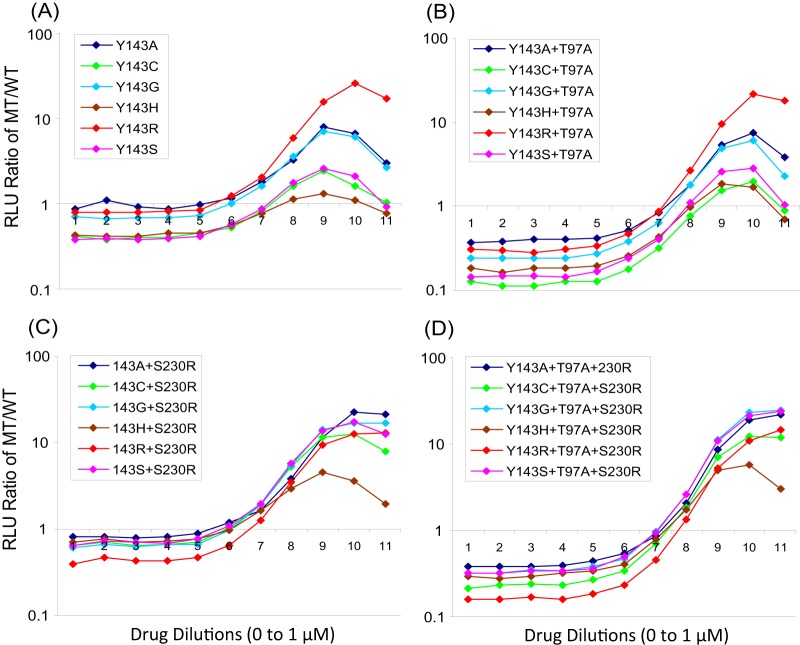
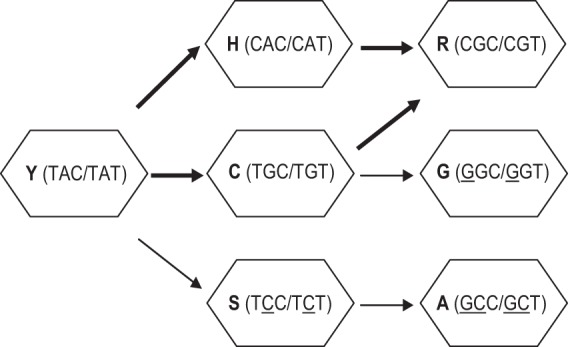
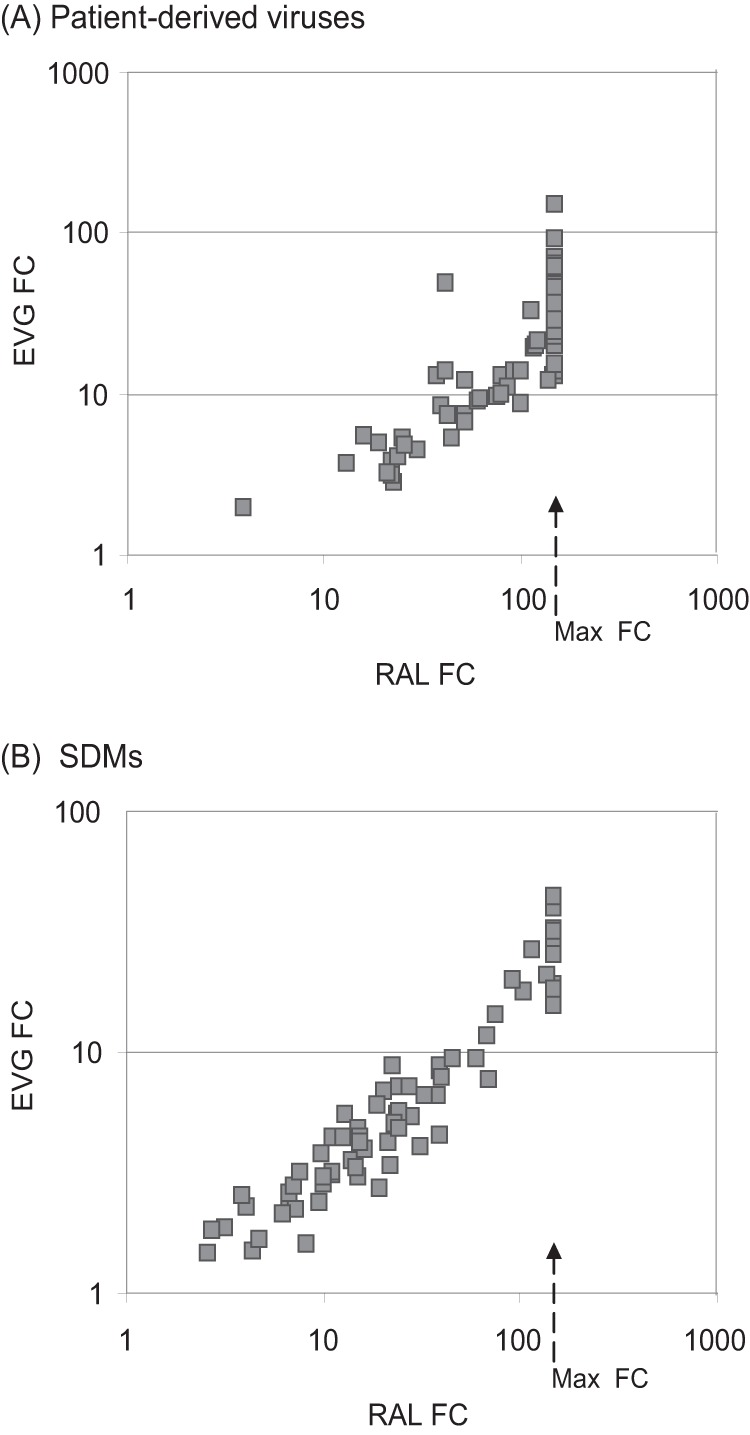
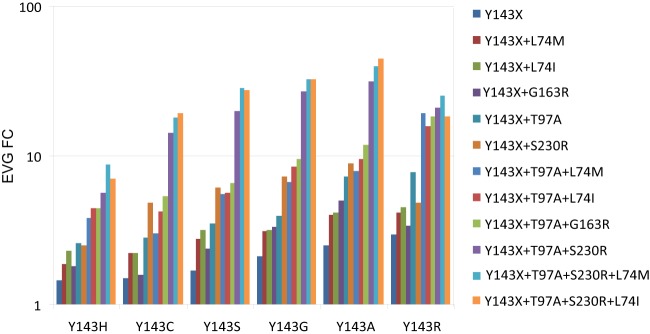
Y143C,R substitutions in HIV-1 integrase define one of three primary raltegravir (RAL) resistance pathways. Here we describe clinical isolates with alternative substitutions at position 143 (Y143A, Y143G, Y143H, and Y143S [Y143A,G,H,S]) that emerge less frequently, and we compare the genotypic and phenotypic profiles of these viruses to Y143C,R viruses to reconcile the preferential selection of Y143C,R variants during RAL treatment. Integrase amino acid sequences and RAL susceptibility were characterized in 117 patient isolates submitted for drug resistance testing and contained Y143 amino acid changes. The influence of specific Y143 substitutions on RAL susceptibility and their preferential association with particular secondary substitutions were further defined by evaluating the composition of patient virus populations along with a large panel of site-directed mutants. Our observations demonstrate that the RAL resistance profiles of Y143A,G,H,S viruses and their association with specific secondary substitutions are similar to the well-established Y143C profile but distinct from the Y143R profile. Y143R viruses differ from Y143A,C,G,H,S viruses in that Y143R confers a greater reduction in RAL susceptibility as a single substitution, consistent with a lower resistance barrier. Among Y143A,C,G,H,S viruses, the higher prevalence of Y143C viruses is the result of a lower genetic barrier than that of the Y143A,G,S viruses and a lower resistance barrier than that of the Y143H viruses. In addition, Y143A,C,G,H,S viruses require multiple secondary substitutions to develop large reductions in RAL susceptibility. Patient-derived viruses containing Y143 substitutions exhibit cross-resistance to elvitegravir.
Figures

References
-
- Pannecouque C, Pluymers W, Van Maele B, Tetz V, Cherepanov P, De Clercq E, Witvrouw M, Debyser Z. 2002. New class of HIV integrase inhibitors that block viral replication in cell culture. Curr. Biol. 12:1169–1177 - PubMed
-
- Bushman FD, Fujiwara T, Craigie R. 1990. Retroviral DNA integration directed by HIV integration protein in vitro. Science 249:1555–1558 - PubMed
-
- Hazuda DJ, Felock P, Witmer M, Wolfe A, Stillmock K, Grobler JA, Espeseth A, Gabryelski L, Schleif W, Blau C, Miller MD. 2000. Inhibitors of strand transfer that prevent integration and inhibit HIV-1 replication in cells. Science 287:646–650 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
