

Fetal-maternal alignment of regulatory T cells correlates with IL-10 and Bcl-2 upregulation in pregnancy
- PMID: 23733877
- PMCID: PMC3690326
- DOI: 10.4049/jimmunol.1203165
Fetal-maternal alignment of regulatory T cells correlates with IL-10 and Bcl-2 upregulation in pregnancy
Abstract
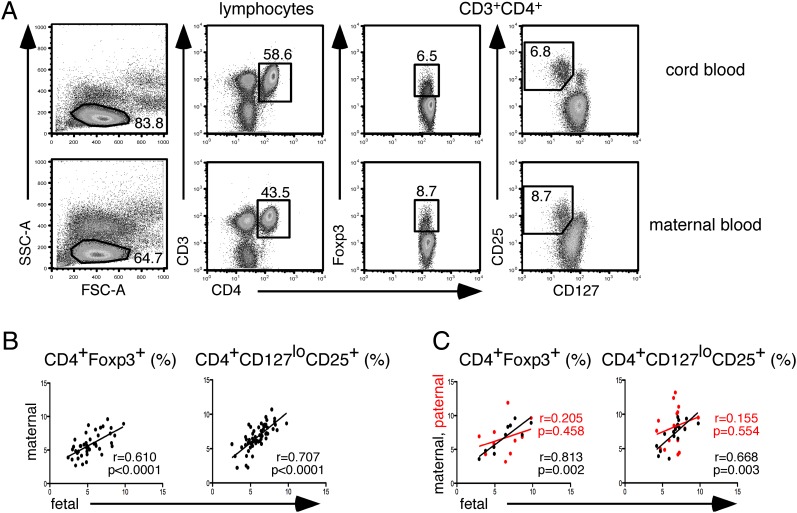
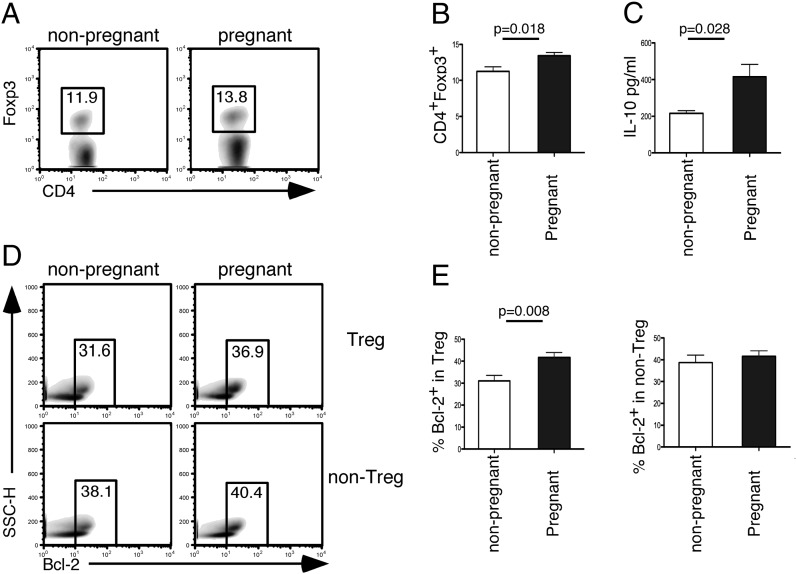
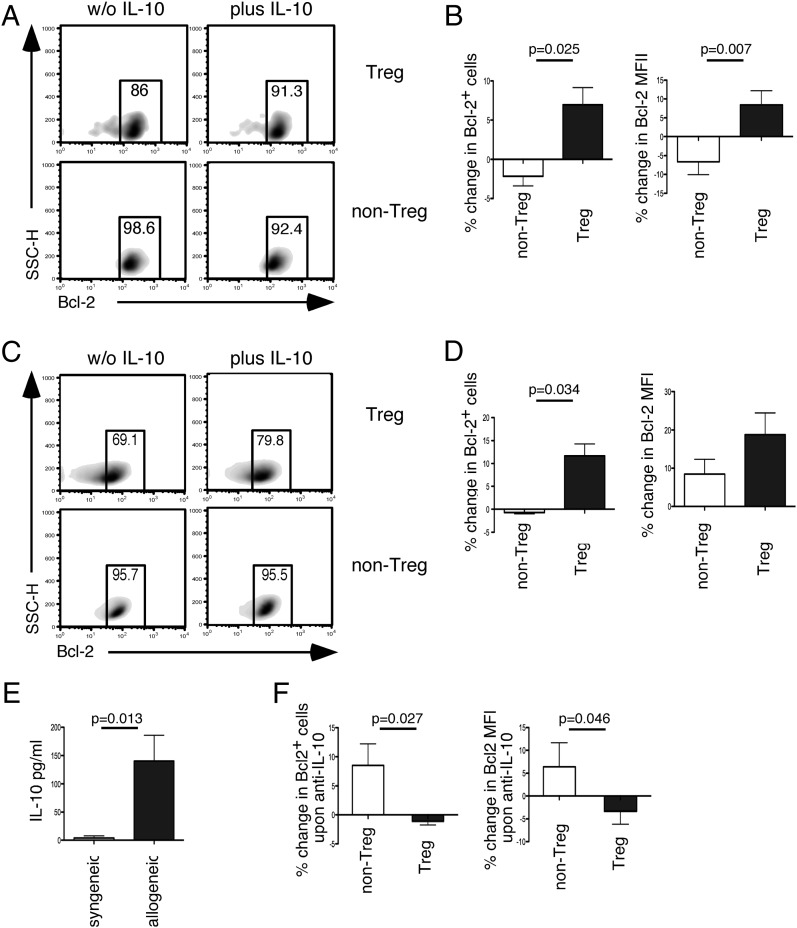
Transplacental immune regulation refers to the concept that during pregnancy, significant cross-talk occurs between the maternal and fetal immune system with potential long-term effects for both the mother and child. In this study, we made the surprising observation that there is a strong correlation of peripheral blood regulatory T (Treg) cells between the mother and the fetus. In contrast, there is no significant Treg cell correlation between paternal fetal dyads (pairs), suggesting that the specific context of pregnancy, rather than the genetic parental similarity to the fetus, is responsible for this correlation. Gene microarray analysis of Treg cells identified a typical IL-10-dependent signature in maternal and fetal Treg cells. In addition, a direct correlation of serum IL-10 protein levels between maternal fetal dyads was observed. Furthermore, we show that maternal serum IL-10 levels correlate with serum estradiol and estriol, implicating hormonal involvement in this alignment. Interestingly, we show that Treg cells possess higher expression of IL-10 receptor α and that Treg cell IL-10 receptor α expression directly correlates with their Bcl-2 expression. Indeed, in vitro data in both humans and mice demonstrate that IL-10 upregulates Bcl-2 specifically in Treg cells but not non-Treg cells. Our results provide evidence for transplacental regulation of cellular immunity and suggest that IL-10 may influence Treg cell homeostasis through its effect on Treg cell Bcl-2 expression. These novel findings have important implications on immune tolerance in pregnancy and beyond in areas of autoimmunity, allergy, and transplantation.
Figures



References
-
- Malassiné A., Frendo J. L., Evain-Brion D. 2003. A comparison of placental development and endocrine functions between the human and mouse model. Hum. Reprod. Update 9: 531–539 - PubMed
-
- Firan M., Bawdon R., Radu C., Ober R. J., Eaken D., Antohe F., Ghetie V., Ward E. S. 2001. The MHC class I-related receptor, FcRn, plays an essential role in the maternofetal transfer of gamma-globulin in humans. Int. Immunol. 13: 993–1002 - PubMed
-
- Loubière L. S., Lambert N. C., Flinn L. J., Erickson T. D., Yan Z., Guthrie K. A., Vickers K. T., Nelson J. L. 2006. Maternal microchimerism in healthy adults in lymphocytes, monocyte/macrophages and NK cells. Lab. Invest. 86: 1185–1192 - PubMed
-
- Macaubas C., de Klerk N. H., Holt B. J., Wee C., Kendall G., Firth M., Sly P. D., Holt P. G. 2003. Association between antenatal cytokine production and the development of atopy and asthma at age 6 years. Lancet 362: 1192–1197 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
