

The diverse functional LINCs of the nuclear envelope to the cytoskeleton and chromatin
- PMID: 23736899
- PMCID: PMC3777164
- DOI: 10.1007/s00412-013-0417-x
The diverse functional LINCs of the nuclear envelope to the cytoskeleton and chromatin
Abstract
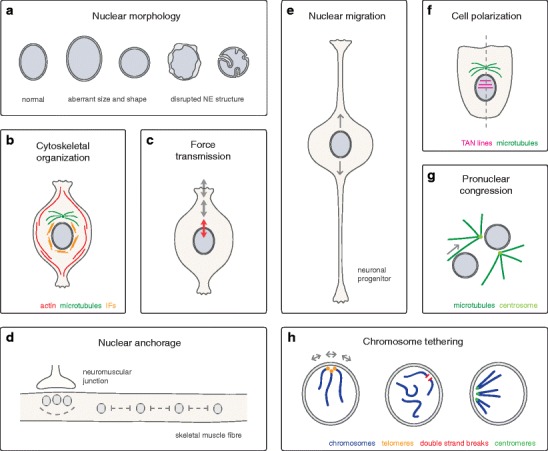
The nuclear envelope (NE) is connected to the different types of cytoskeletal elements by linker of nucleoskeleton and cytoskeleton (LINC) complexes. LINC complexes exist from yeast to humans, and have preserved their general architecture throughout evolution. They are composed of SUN and KASH domain proteins of the inner and the outer nuclear membrane, respectively. These SUN-KASH bridges are used for the transmission of forces across the NE and support diverse biological processes. Here, we review the function of SUN and KASH domain proteins in various unicellular and multicellular species. Specifically, we discuss their influence on nuclear morphology and cytoskeletal organization. Further, emphasis is given on the role of LINC complexes in nuclear anchorage and migration as well as in genome organization.
Figures

References
-
- Behrens TW, Jagadeesh J, Scherle P, Kearns G, Yewdell J, Staudt LM. Jaw1, A lymphoid-restricted membrane protein localized to the endoplasmic reticulum. J Immunol. 1994;153(2):682–690. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
