

The chlorite dismutase (HemQ) from Staphylococcus aureus has a redox-sensitive heme and is associated with the small colony variant phenotype
- PMID: 23737523
- PMCID: PMC5395028
- DOI: 10.1074/jbc.M112.442335
The chlorite dismutase (HemQ) from Staphylococcus aureus has a redox-sensitive heme and is associated with the small colony variant phenotype
Abstract
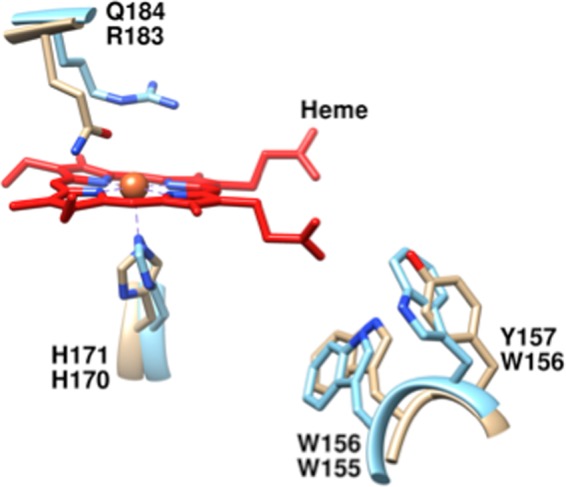
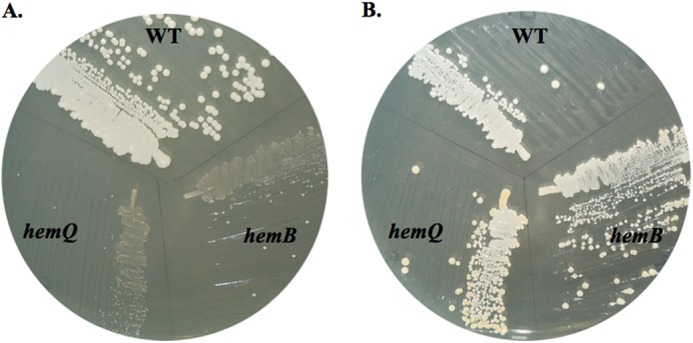
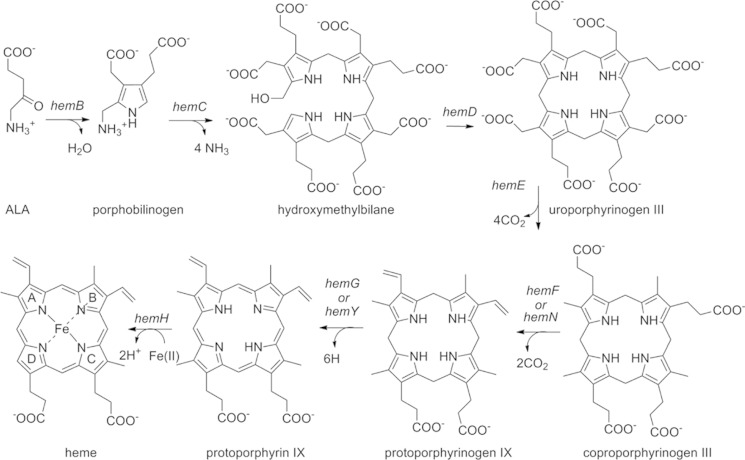
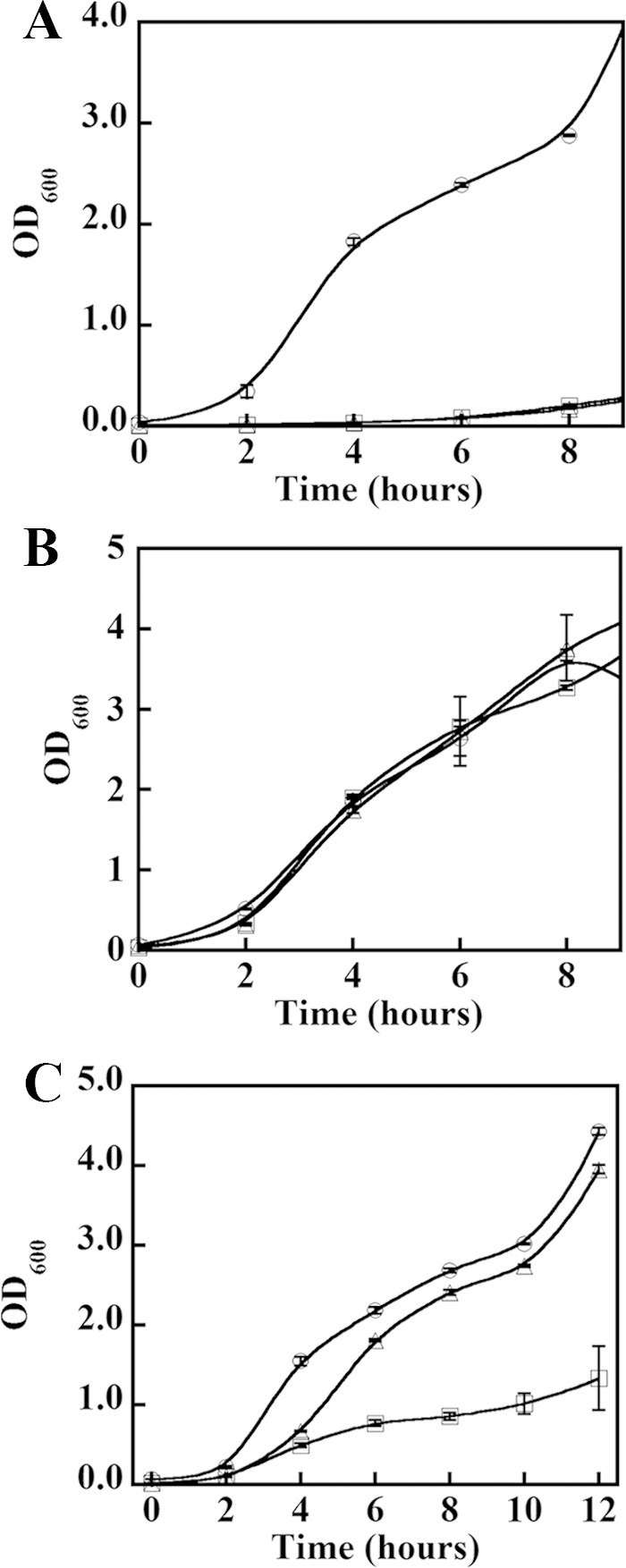
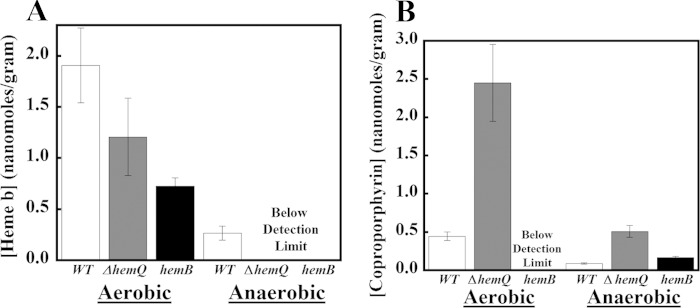
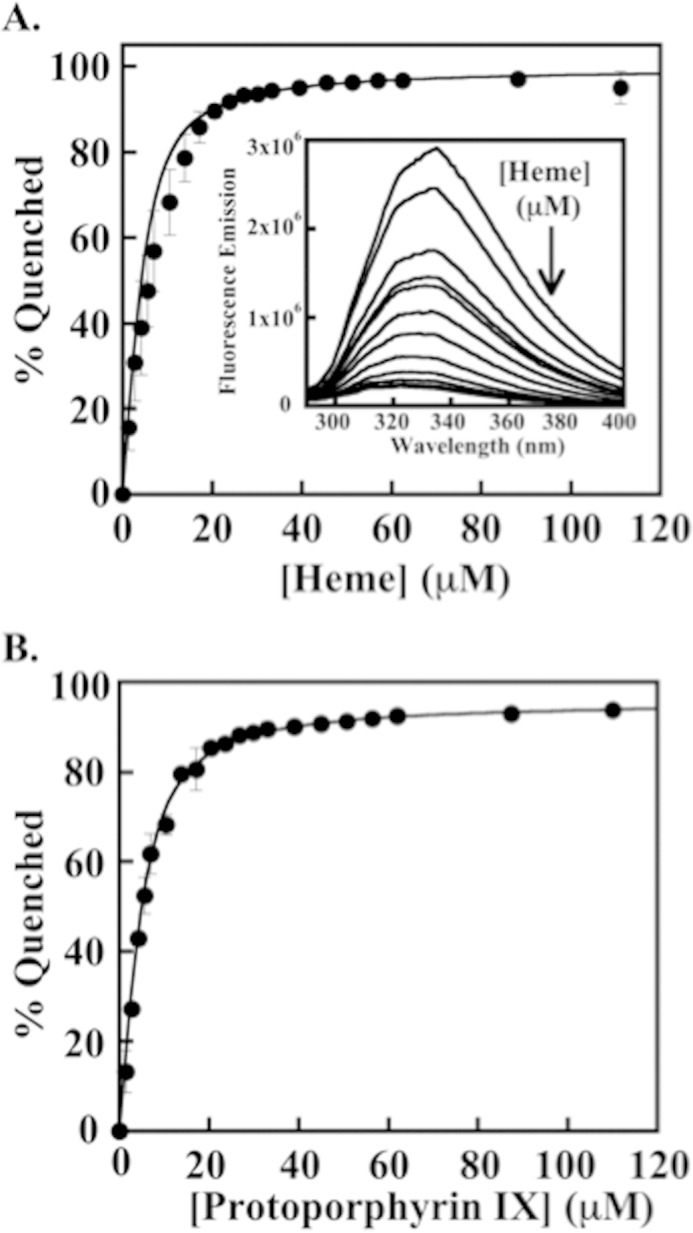
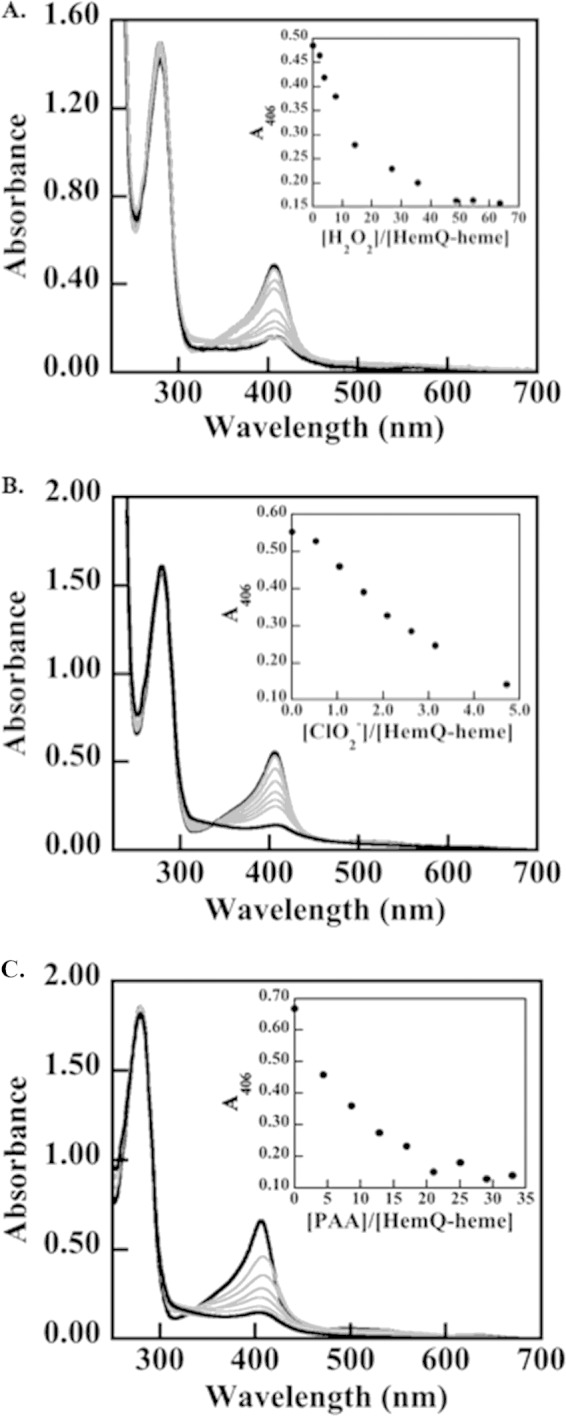
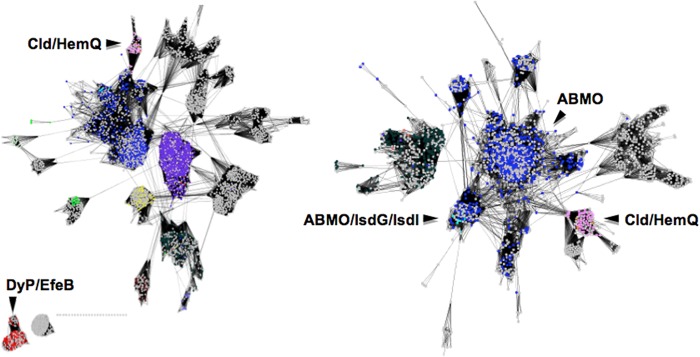
The chlorite dismutases (C-family proteins) are a widespread family of heme-binding proteins for which chemical and biological roles remain unclear. An association of the gene with heme biosynthesis in Gram-positive bacteria was previously demonstrated by experiments involving introduction of genes from two Gram-positive species into heme biosynthesis mutant strains of Escherichia coli, leading to the gene being renamed hemQ. To assess the gene product's biological role more directly, a Staphylococcus aureus strain with an inactivated hemQ gene was generated and shown to be a slow growing small colony variant under aerobic but not anaerobic conditions. The small colony variant phenotype is rescued by the addition of exogenous heme despite an otherwise wild type heme biosynthetic pathway. The ΔhemQ mutant accumulates coproporphyrin specifically under aerobic conditions. Although its sequence is highly similar to functional chlorite dismutases, the HemQ protein has no steady state reactivity with chlorite, very modest reactivity with H2O2 or peracetic acid, and no observable transient intermediates. HemQ's equilibrium affinity for heme is in the low micromolar range. Holo-HemQ reconstituted with heme exhibits heme lysis after <50 turnovers with peroxide and <10 turnovers with chlorite. The heme-free apoprotein aggregates or unfolds over time. IsdG-like proteins and antibiotic biosynthesis monooxygenases are close sequence and structural relatives of HemQ that use heme or porphyrin-like organic molecules as substrates. The genetic and biochemical data suggest a similar substrate role for heme or porphyrin, with possible sensor-regulator functions for the protein. HemQ heme could serve as the means by which S. aureus reversibly adopts an SCV phenotype in response to redox stress.
Keywords: Bacteria; Chlorite Dismutase; Heme; Peroxidase; Redox Regulation; Staphylococcus aureus; Superoxide Ion.
Figures
References
-
- Coates J. D., Achenbach L. A. (2004) Microbial perchlorate reduction: Rocket-fuelled metabolism. Nat. Rev. Microbiol. 2, 569–580 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Chemical Information
