

Bone morphogenetic protein 9 (BMP9) controls lymphatic vessel maturation and valve formation
- PMID: 23741013
- PMCID: PMC3724195
- DOI: 10.1182/blood-2012-12-472142
Bone morphogenetic protein 9 (BMP9) controls lymphatic vessel maturation and valve formation
Abstract
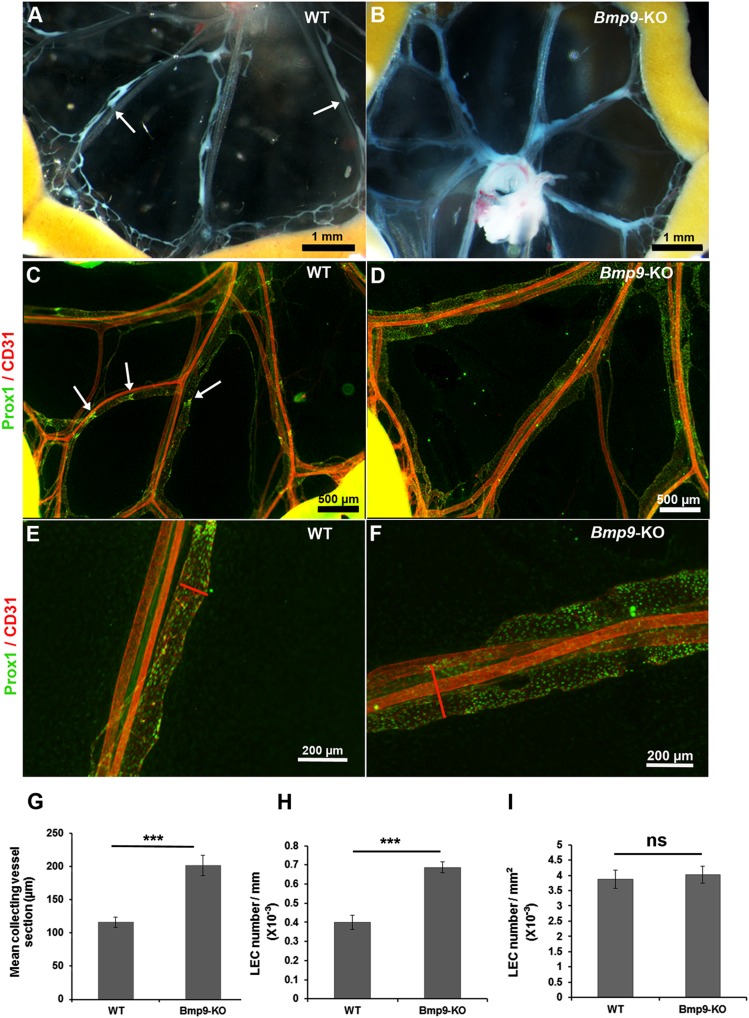
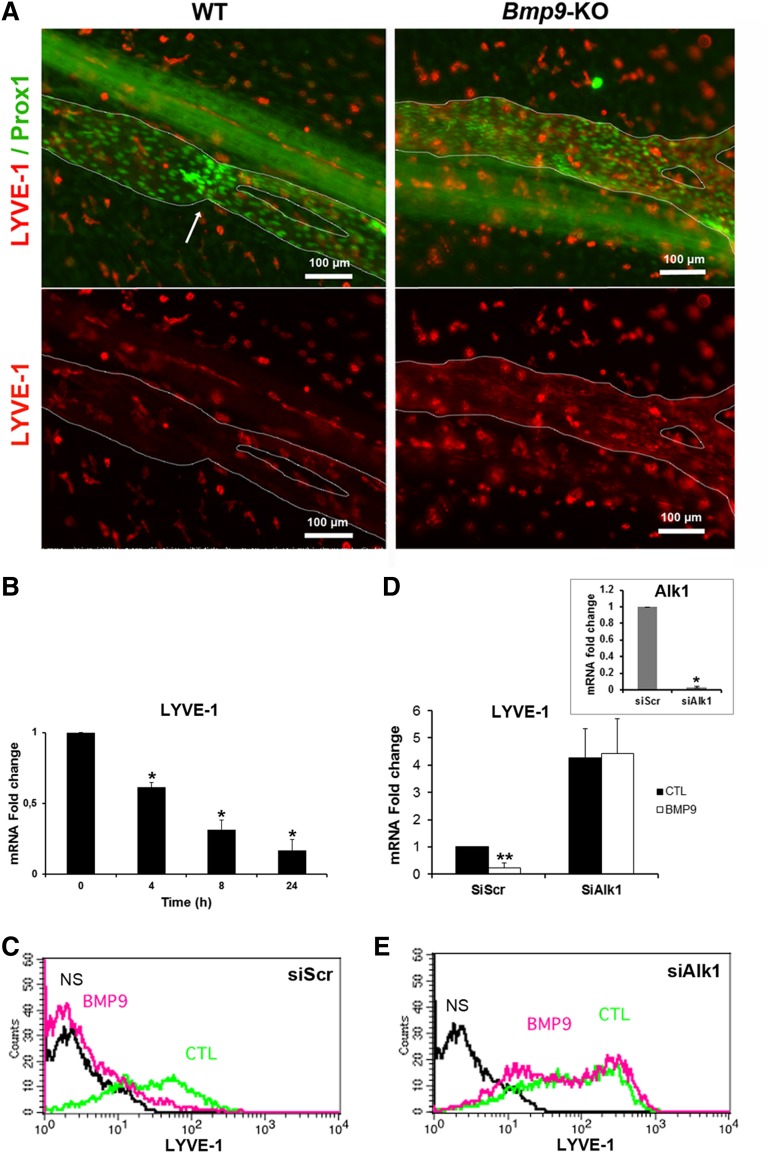
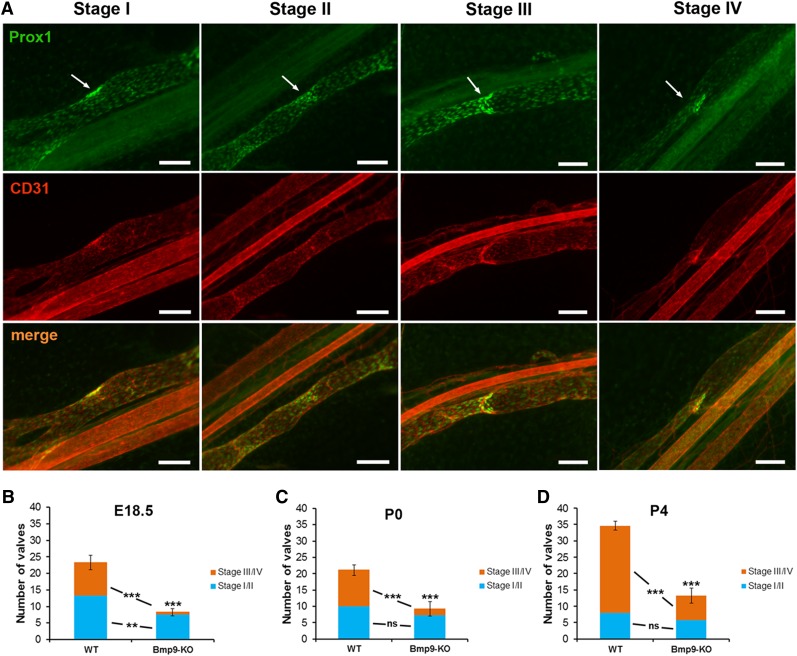
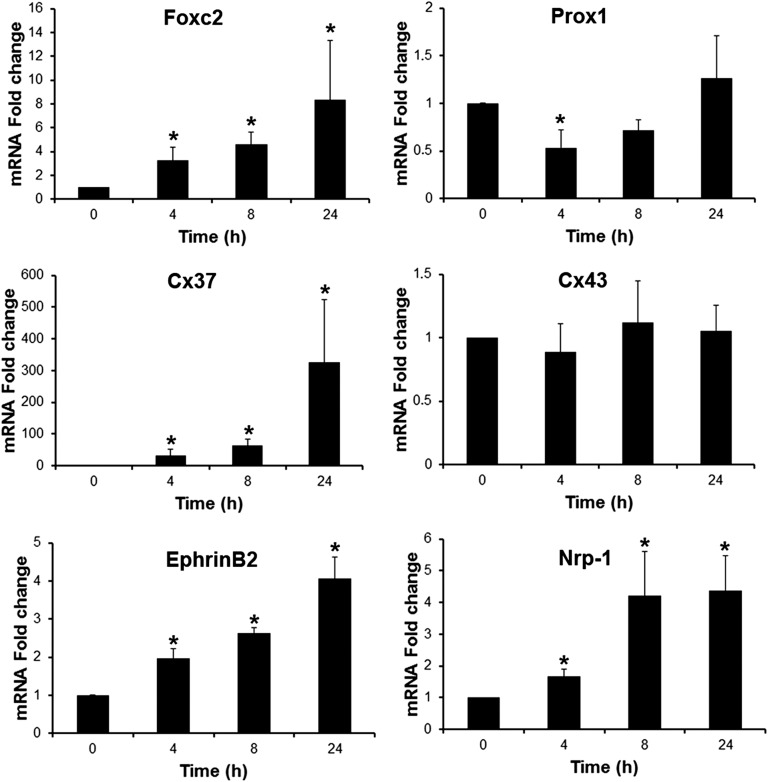
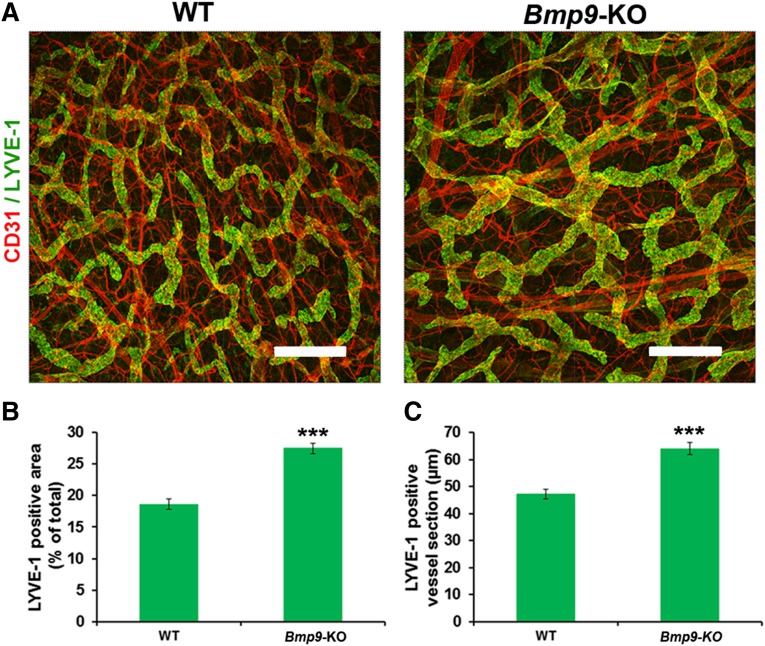
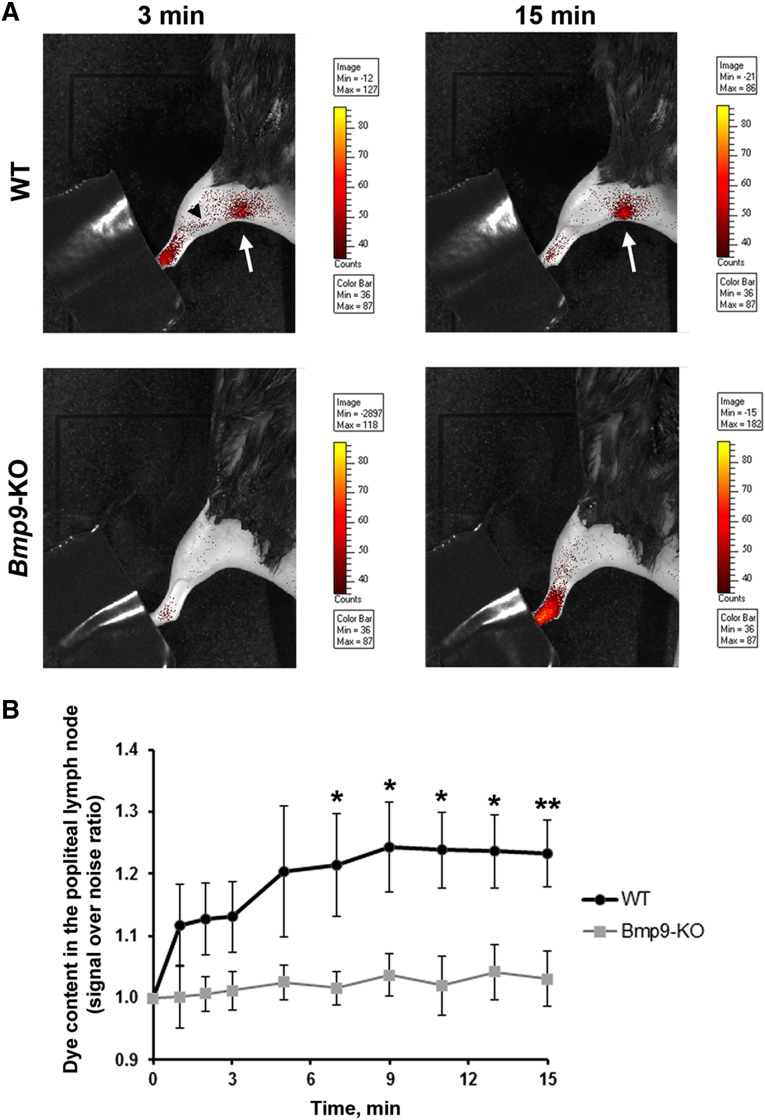
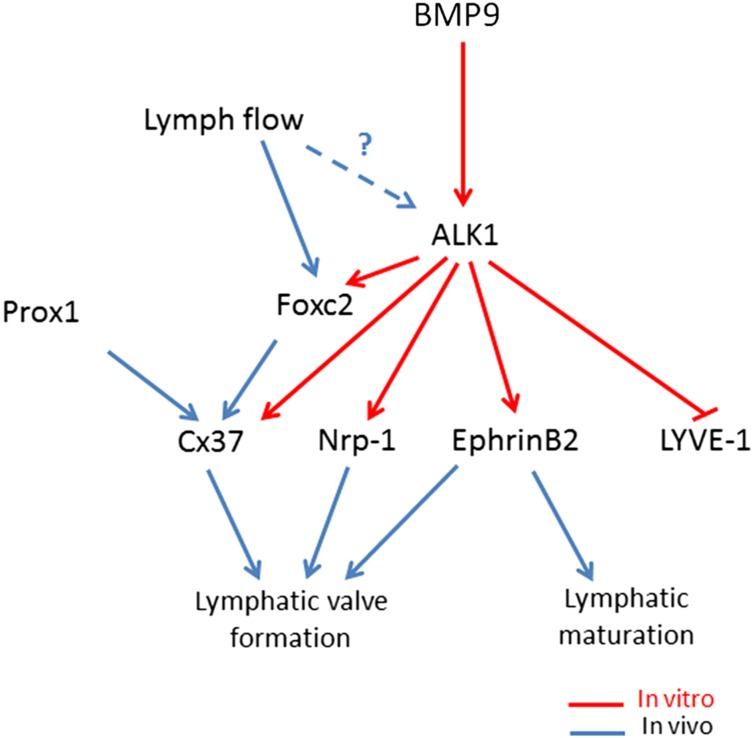
Lymphatic vessels are critical for the maintenance of tissue fluid homeostasis and their dysfunction contributes to several human diseases. The activin receptor-like kinase 1 (ALK1) is a transforming growth factor-β family type 1 receptor that is expressed on both blood and lymphatic endothelial cells (LECs). Its high-affinity ligand, bone morphogenetic protein 9 (BMP9), has been shown to be critical for retinal angiogenesis. The aim of this work was to investigate whether BMP9 could play a role in lymphatic development. We found that Bmp9 deficiency in mice causes abnormal lymphatic development. Bmp9-knockout (KO) pups presented hyperplastic mesenteric collecting vessels that maintained LYVE-1 expression. In accordance with this result, we found that BMP9 inhibited LYVE-1 expression in LECs in an ALK1-dependent manner. Bmp9-KO pups also presented a significant reduction in the number and in the maturation of mesenteric lymphatic valves at embryonic day 18.5 and at postnatal days 0 and 4. Interestingly, the expression of several genes known to be involved in valve formation (Foxc2, Connexin37, EphrinB2, and Neuropilin1) was upregulated by BMP9 in LECS. Finally, we demonstrated that Bmp9-KO neonates and adult mice had decreased lymphatic draining efficiency. These data identify BMP9 as an important extracellular regulator in the maturation of the lymphatic vascular network affecting valve development and lymphatic vessel function.
Figures
Similar articles
-
BMP9 and BMP10 are critical for postnatal retinal vascular remodeling.Blood. 2012 Jun 21;119(25):6162-71. doi: 10.1182/blood-2012-01-407593. Epub 2012 May 7. Blood. 2012. PMID: 22566602 Free PMC article.
-
Eye lymphatic defects induced by bone morphogenetic protein 9 deficiency have no functional consequences on intraocular pressure.Sci Rep. 2020 Sep 29;10(1):16040. doi: 10.1038/s41598-020-71877-z. Sci Rep. 2020. PMID: 32994463 Free PMC article.
-
Bone Morphogenetic Protein 9 Is a Paracrine Factor Controlling Liver Sinusoidal Endothelial Cell Fenestration and Protecting Against Hepatic Fibrosis.Hepatology. 2019 Oct;70(4):1392-1408. doi: 10.1002/hep.30655. Epub 2019 May 31. Hepatology. 2019. PMID: 30964206
-
BMP9 and BMP10: Two close vascular quiescence partners that stand out.Dev Dyn. 2022 Jan;251(1):178-197. doi: 10.1002/dvdy.395. Epub 2021 Jul 19. Dev Dyn. 2022. PMID: 34240497 Review.
-
Lymphatic collecting vessel maturation and valve morphogenesis.Microvasc Res. 2014 Nov;96:31-7. doi: 10.1016/j.mvr.2014.07.001. Epub 2014 Jul 12. Microvasc Res. 2014. PMID: 25020266 Review.
Cited by
-
The endometrial transcriptome transition preceding receptivity to embryo implantation in mice.BMC Genomics. 2023 Oct 4;24(1):590. doi: 10.1186/s12864-023-09698-3. BMC Genomics. 2023. PMID: 37794337 Free PMC article.
-
A heterodimer formed by bone morphogenetic protein 9 (BMP9) and BMP10 provides most BMP biological activity in plasma.J Biol Chem. 2018 Jul 13;293(28):10963-10974. doi: 10.1074/jbc.RA118.002968. Epub 2018 May 22. J Biol Chem. 2018. PMID: 29789425 Free PMC article.
-
Endothelial depletion of Acvrl1 in mice leads to arteriovenous malformations associated with reduced endoglin expression.PLoS One. 2014 Jun 4;9(6):e98646. doi: 10.1371/journal.pone.0098646. eCollection 2014. PLoS One. 2014. PMID: 24896812 Free PMC article.
-
Mechanosensation and Mechanotransduction by Lymphatic Endothelial Cells Act as Important Regulators of Lymphatic Development and Function.Int J Mol Sci. 2021 Apr 12;22(8):3955. doi: 10.3390/ijms22083955. Int J Mol Sci. 2021. PMID: 33921229 Free PMC article. Review.
-
Tie1 is required for lymphatic valve and collecting vessel development.Dev Biol. 2015 Mar 1;399(1):117-128. doi: 10.1016/j.ydbio.2014.12.021. Epub 2015 Jan 7. Dev Biol. 2015. PMID: 25576926 Free PMC article.
References
-
- Tammela T, Alitalo K. Lymphangiogenesis: molecular mechanisms and future promise. Cell. 2010;140(4):460–476. - PubMed
-
- Rockson SG. Diagnosis and management of lymphatic vascular disease. J Am Coll Cardiol. 2008;52(10):799–806. - PubMed
-
- Norrmén C, Tammela T, Petrova TV, Alitalo K. Biological basis of therapeutic lymphangiogenesis. Circulation. 2011;123(12):1335–1351. - PubMed
-
- Petrova TV, Karpanen T, Norrmén C, et al. Defective valves and abnormal mural cell recruitment underlie lymphatic vascular failure in lymphedema distichiasis. Nat Med. 2004;10(9):974–981. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
