

An MLL-dependent network sustains hematopoiesis
- PMID: 23744037
- PMCID: PMC3718146
- DOI: 10.1073/pnas.1301278110
An MLL-dependent network sustains hematopoiesis
Abstract
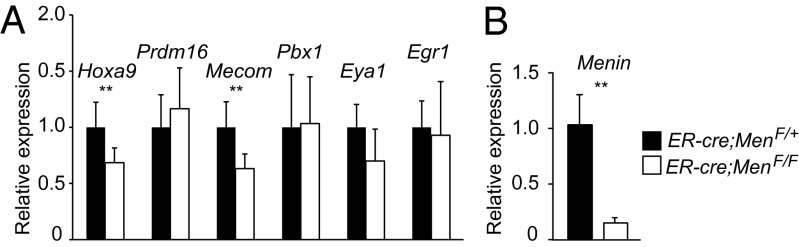
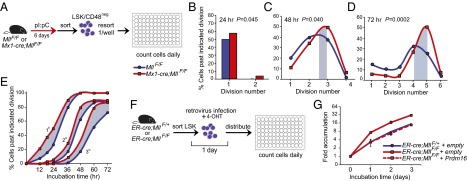
The histone methyltransferase Mixed Lineage Leukemia (MLL) is essential to maintain hematopoietic stem cells and is a leukemia protooncogene. Although clustered homeobox genes are well-characterized targets of MLL and MLL fusion oncoproteins, the range of Mll-regulated genes in normal hematopoietic cells remains unknown. Here, we identify and characterize part of the Mll-dependent transcriptional network in hematopoietic stem cells with an integrated approach by using conditional loss-of-function models, genomewide expression analyses, chromatin immunoprecipitation, and functional rescue assays. The Mll-dependent transcriptional network extends well beyond the previously appreciated Hox targets, is comprised of many characterized regulators of self-renewal, and contains target genes that are both dependent and independent of the MLL cofactor, Menin. Interestingly, PR-domain containing 16 emerged as a target gene that is uniquely effective at partially rescuing Mll-deficient hematopoietic stem and progenitor cells. This work highlights the tissue-specific nature of regulatory networks under the control of MLL/Trithorax family members and provides insight into the distinctions between the participation of MLL in normal hematopoiesis and in leukemia.
Keywords: HSC; epigenetics; proliferation.
Conflict of interest statement
Conflict of interest statement: P.E. is a shareholder of Amgen stock.
Figures




Comment in
-
Delineating the mixed lineage leukemia gene expression network in hematopoietic stem cells.Proc Natl Acad Sci U S A. 2013 Jul 16;110(29):11670-1. doi: 10.1073/pnas.1310349110. Epub 2013 Jul 3. Proc Natl Acad Sci U S A. 2013. PMID: 23824294 Free PMC article. No abstract available.
References
-
- Schuettengruber B, Chourrout D, Vervoort M, Leblanc B, Cavalli G. Genome regulation by polycomb and trithorax proteins. Cell. 2007;128(4):735–745. - PubMed
-
- Sparmann A, van Lohuizen M. Polycomb silencers control cell fate, development and cancer. Nat Rev Cancer. 2006;6(11):846–856. - PubMed
-
- Daser A, Rabbitts TH. Extending the repertoire of the mixed-lineage leukemia gene MLL in leukemogenesis. Genes Dev. 2004;18(9):965–974. - PubMed
-
- Krivtsov AV, Armstrong SA. MLL translocations, histone modifications and leukaemia stem-cell development. Nat Rev Cancer. 2007;7(11):823–833. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
