

Lipid, detergent, and Coomassie Blue G-250 affect the migration of small membrane proteins in blue native gels: mitochondrial carriers migrate as monomers not dimers
- PMID: 23744064
- PMCID: PMC3724668
- DOI: 10.1074/jbc.M113.484329
Lipid, detergent, and Coomassie Blue G-250 affect the migration of small membrane proteins in blue native gels: mitochondrial carriers migrate as monomers not dimers
Abstract
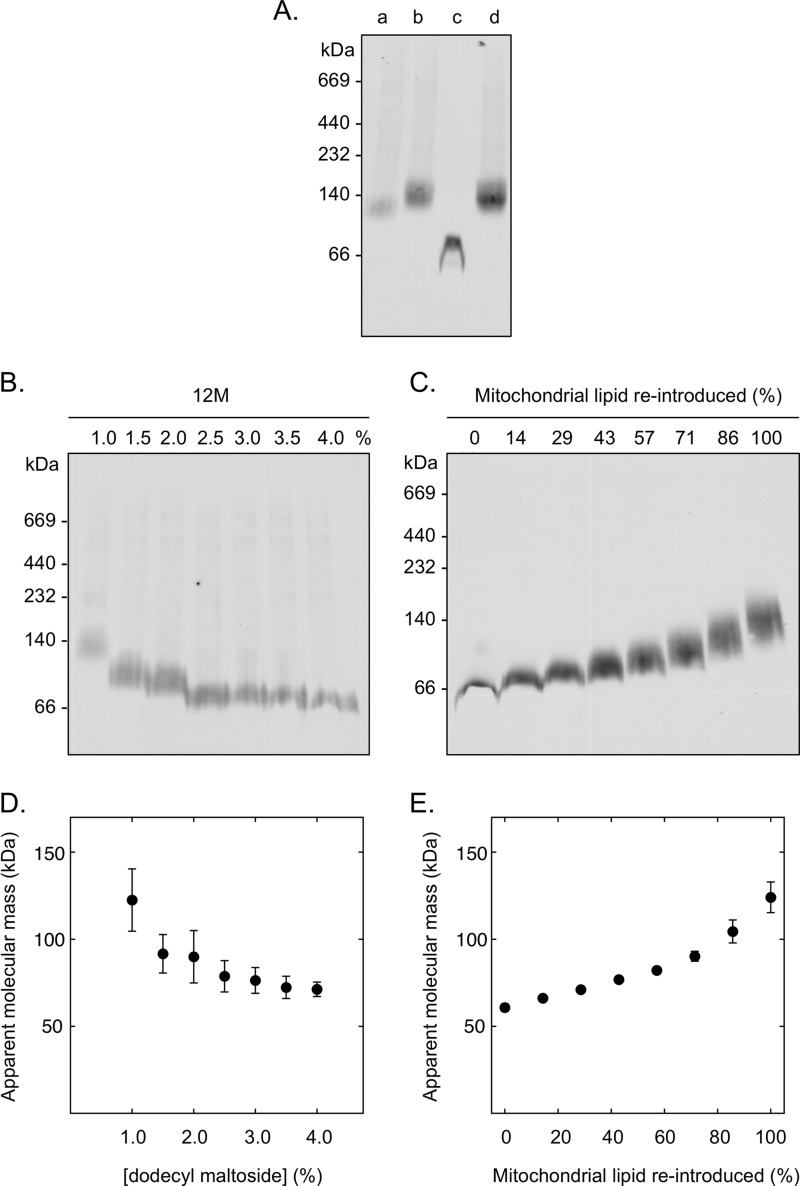
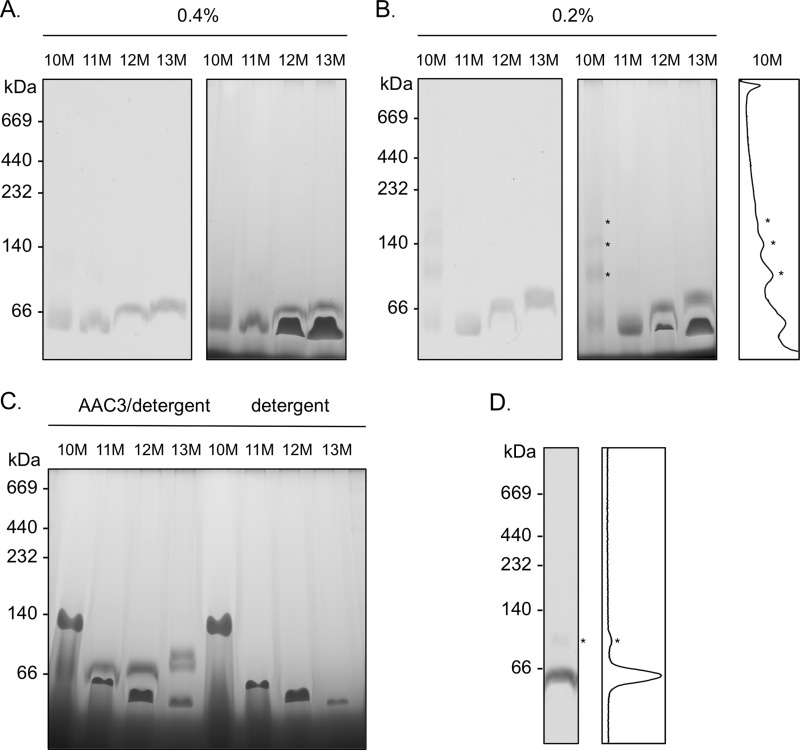
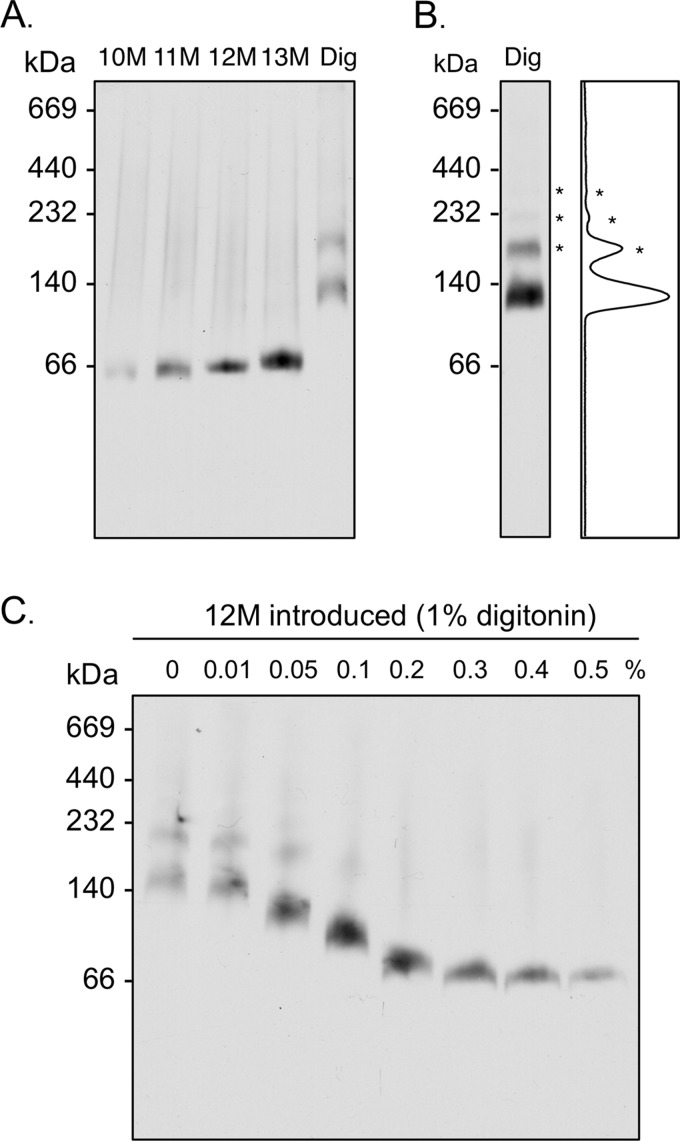
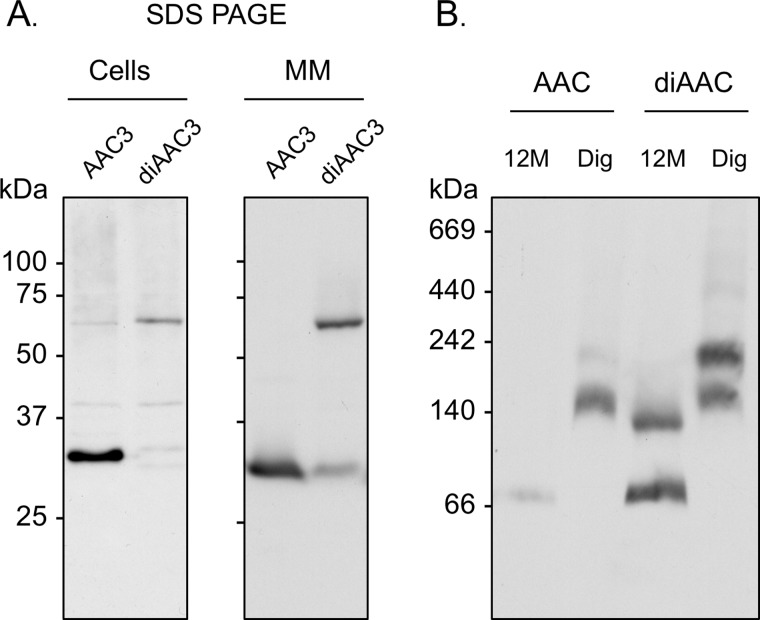
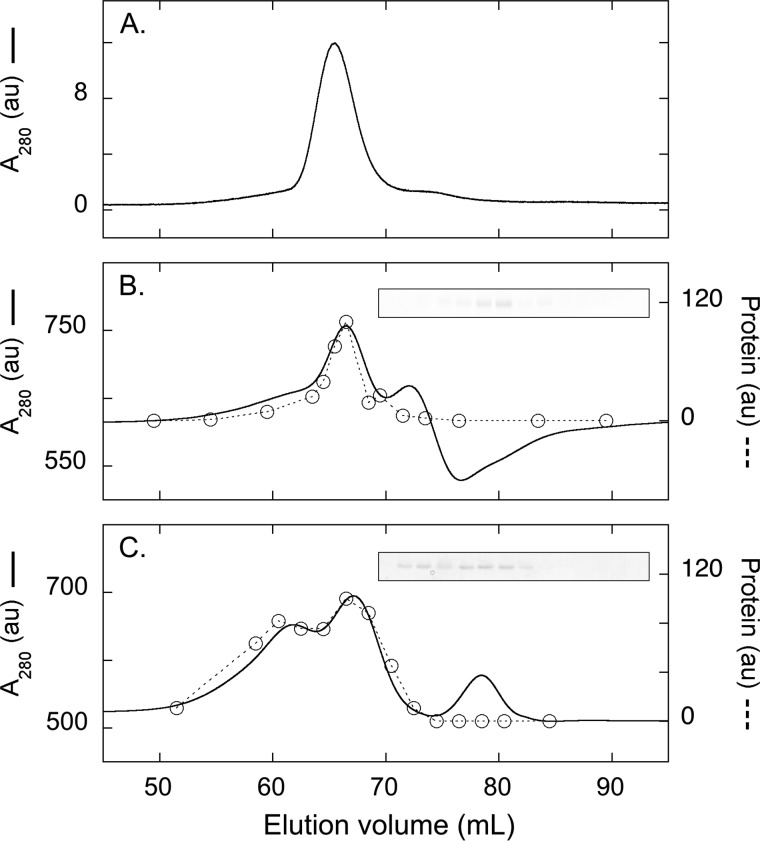
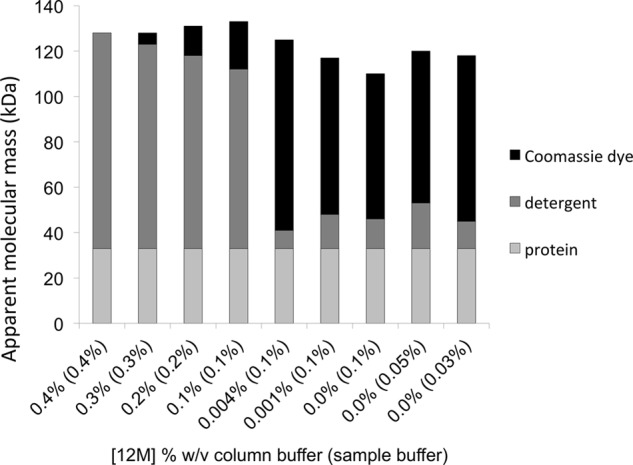
Blue native gel electrophoresis is a popular method for the determination of the oligomeric state of membrane proteins. Studies using this technique have reported that mitochondrial carriers are dimeric (composed of two ∼32-kDa monomers) and, in some cases, can form physiologically relevant associations with other proteins. Here, we have scrutinized the behavior of the yeast mitochondrial ADP/ATP carrier AAC3 in blue native gels. We find that the apparent mass of AAC3 varies in a detergent- and lipid-dependent manner (from ∼60 to ∼130 kDa) that is not related to changes in the oligomeric state of the protein, but reflects differences in the associated detergent-lipid micelle and Coomassie Blue G-250 used in this technique. Higher oligomeric state species are only observed under less favorable solubilization conditions, consistent with aggregation of the protein. Calibration with an artificial covalent AAC3 dimer indicates that the mass observed for solubilized AAC3 and other mitochondrial carriers corresponds to a monomer. Size exclusion chromatography of purified AAC3 in dodecyl maltoside under blue native gel-like conditions shows that the mass of the monomer is ∼120 kDa, but appears smaller on gels (∼60 kDa) due to the unusually high amount of bound negatively charged dye, which increases the electrophoretic mobility of the protein-detergent-dye micelle complex. Our results show that bound lipid, detergent, and Coomassie stain alter the behavior of mitochondrial carriers on gels, which is likely to be true for other small membrane proteins where the associated lipid-detergent micelle is large when compared with the mass of the protein.
Keywords: Detergent Micelle; Gel Electrophoresis; Membrane Proteins; Mitochondrial Transport; Protein Aggregation; Transporters.
Figures

References
-
- Kunji E. R. (2004) The role and structure of mitochondrial carriers. FEBS Lett. 564, 239–244 - PubMed
-
- Palmieri F. (2008) Diseases caused by defects of mitochondrial carriers: a review. Biochim. Biophys. Acta 1777, 564–578 - PubMed
-
- Nedergaard J., Cannon B. (2010) The changed metabolic world with human brown adipose tissue: therapeutic visions. Cell Metab. 11, 268–272 - PubMed
-
- Pebay-Peyroula E., Dahout-Gonzalez C., Kahn R., Trézéguet V., Lauquin G. J., Brandolin G. (2003) Structure of mitochondrial ADP/ATP carrier in complex with carboxyatractyloside. Nature 426, 39–44 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
