

PARG dysfunction enhances DNA double strand break formation in S-phase after alkylation DNA damage and augments different cell death pathways
- PMID: 23744356
- PMCID: PMC3698538
- DOI: 10.1038/cddis.2013.133
PARG dysfunction enhances DNA double strand break formation in S-phase after alkylation DNA damage and augments different cell death pathways
Abstract
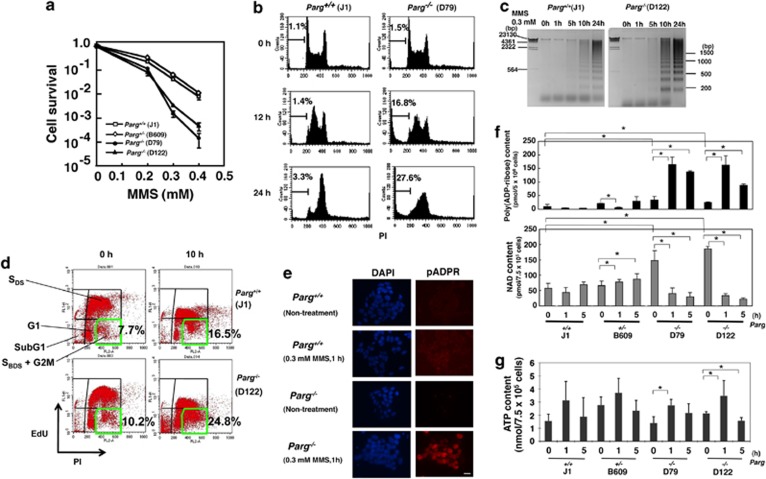
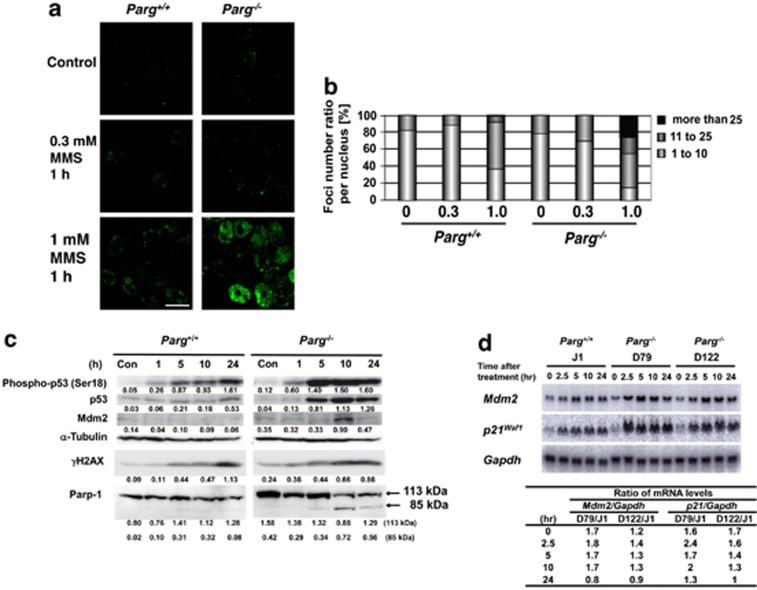
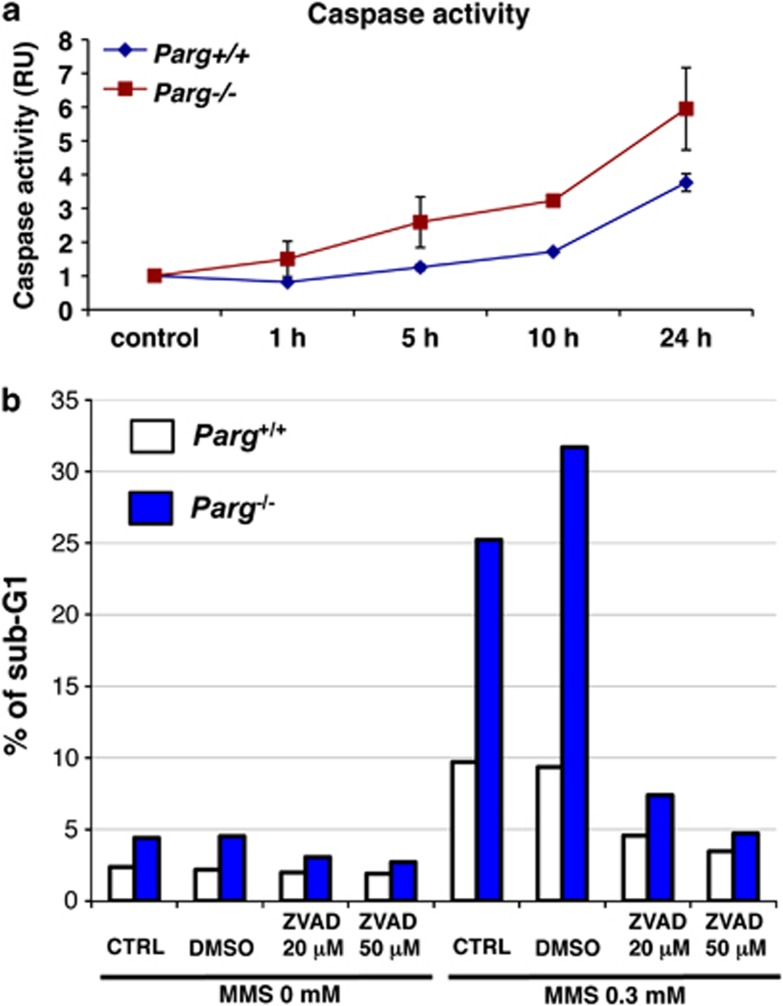
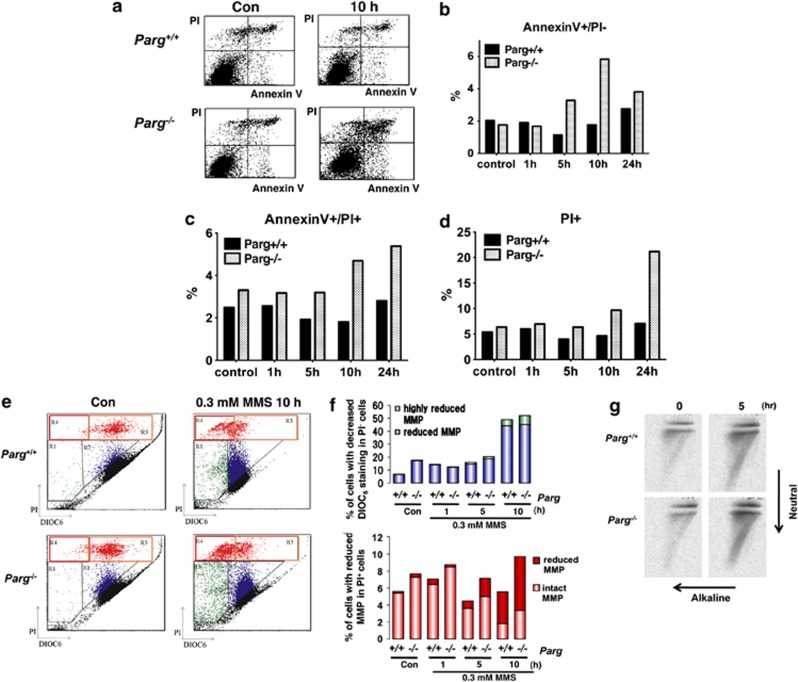
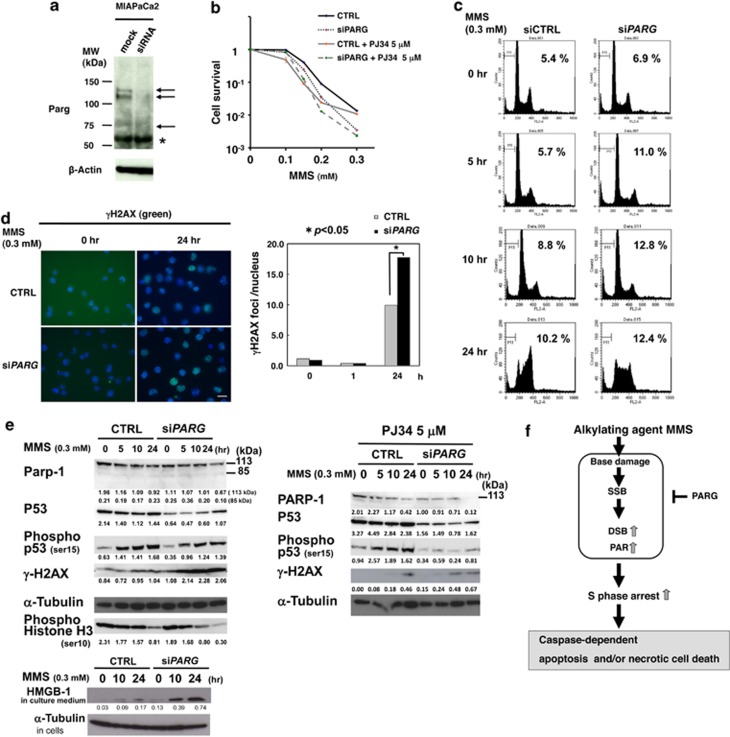
Poly(ADP-ribose) glycohydrolase (PARG) is the primary enzyme responsible for the degradation of poly(ADP-ribose). PARG dysfunction sensitizes cells to alkylating agents and induces cell death; however, the details of this effect have not been fully elucidated. Here, we investigated the mechanism by which PARG deficiency leads to cell death in different cell types using methylmethanesulfonate (MMS), an alkylating agent, and Parg(-/-) mouse ES cells and human cancer cell lines. Parg(-/-) mouse ES cells showed increased levels of γ-H2AX, a marker of DNA double strand breaks (DSBs), accumulation of poly(ADP-ribose), p53 network activation, and S-phase arrest. Early apoptosis was enhanced in Parg(-/-) mouse ES cells. Parg(-/-) ES cells predominantly underwent caspase-dependent apoptosis. PARG was then knocked down in a p53-defective cell line, MIAPaCa2 cells, a human pancreatic cancer cell line. MIAPaCa2 cells were sensitized to MMS by PARG knockdown. Enhanced necrotic cell death was induced in MIAPaCa2 cells after augmenting γ-H2AX levels and S-phase arrest. Taken together, these data suggest that DSB repair defect causing S-phase arrest, but p53 status was not important for sensitization to alkylation DNA damage by PARG dysfunction, whereas the cell death pathway is dependent on the cell type. This study demonstrates that functional inhibition of PARG may be useful for sensitizing at least particular cancer cells to alkylating agents.
Figures
References
-
- Sugimura T. Poly(adenosine diphosphate ribose) Prog Nucleic Acid Res Mol Biol. 1973;13:127–151. - PubMed
-
- Meyer-Ficca ML, Meyer RG, Coyle DL, Jacobson EL, Jacobson MK. Human poly(ADP-ribose) glycohydrolase is expressed in alternative splice variants yielding isoforms that localize to different cell compartments. Exp Cell Res. 2004;297:521–532. - PubMed
-
- Fujihara H, Ogino H, Maeda D, Shirai H, Nozaki T, Kamada N, et al. Poly(ADP-ribose) Glycohydrolase deficiency sensitizes mouse ES cells to DNA damaging agents. Curr Cancer Drug Targets. 2009;9:953–962. - PubMed
-
- Oka S, Kato J, Moss J. Identification and characterization of a mammalian 39-kDa poly(ADP-ribose) glycohydrolase. J Biol Chem. 2006;281:705–713. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
