

Posttranscriptional control of T cell effector function by aerobic glycolysis
- PMID: 23746840
- PMCID: PMC3804311
- DOI: 10.1016/j.cell.2013.05.016
Posttranscriptional control of T cell effector function by aerobic glycolysis
Abstract
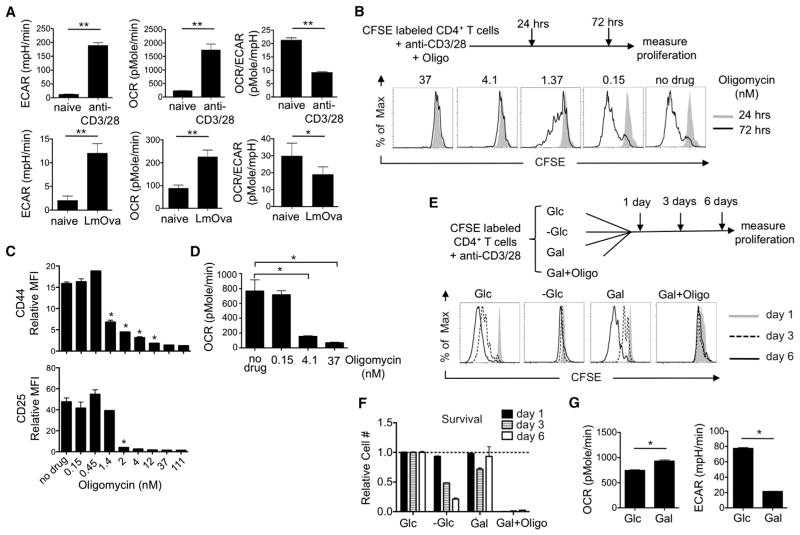
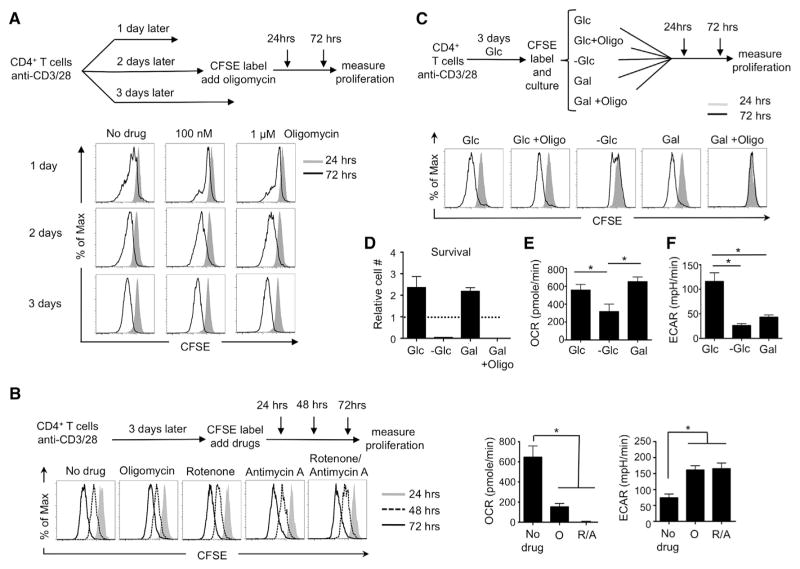
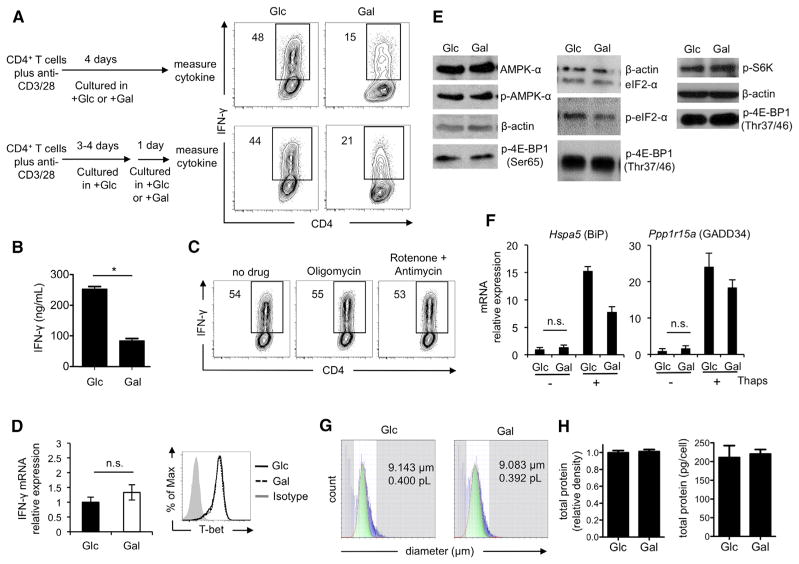
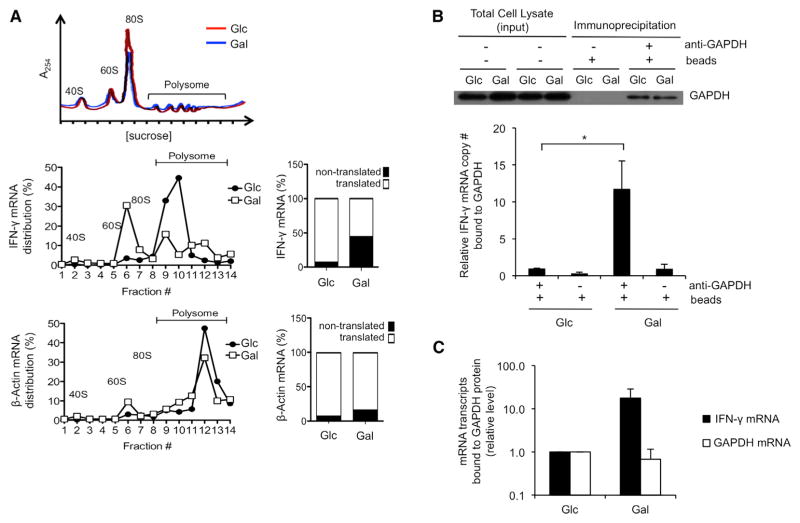
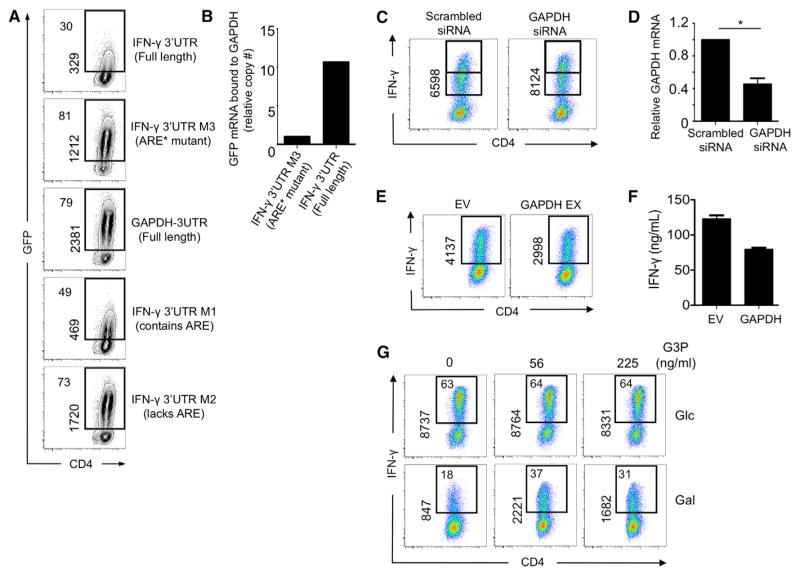
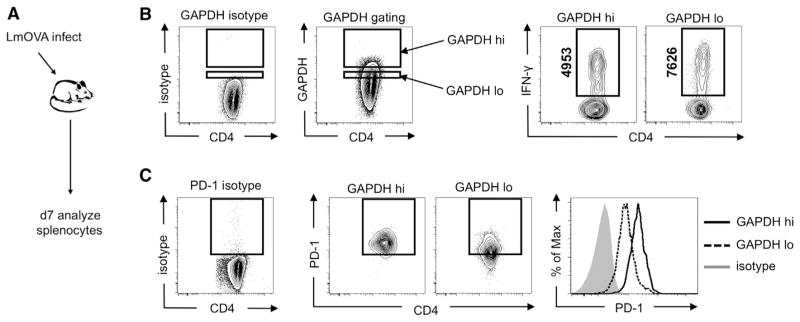
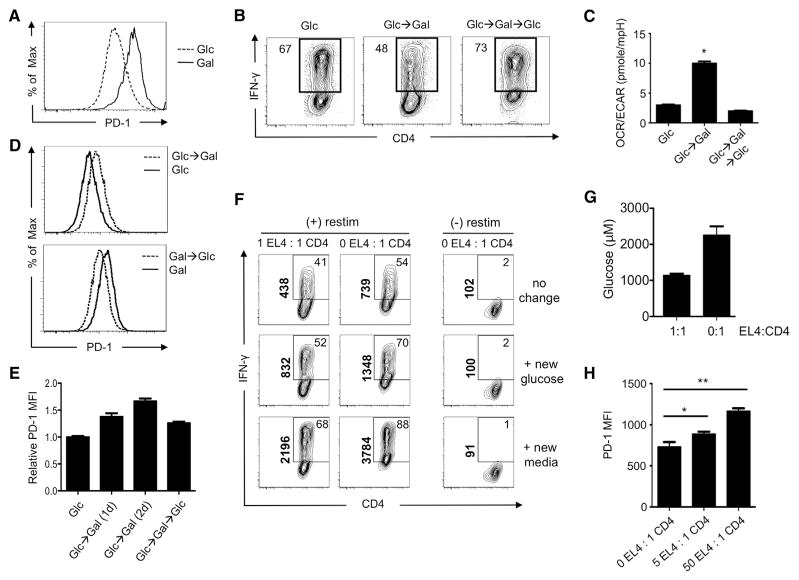
A "switch" from oxidative phosphorylation (OXPHOS) to aerobic glycolysis is a hallmark of T cell activation and is thought to be required to meet the metabolic demands of proliferation. However, why proliferating cells adopt this less efficient metabolism, especially in an oxygen-replete environment, remains incompletely understood. We show here that aerobic glycolysis is specifically required for effector function in T cells but that this pathway is not necessary for proliferation or survival. When activated T cells are provided with costimulation and growth factors but are blocked from engaging glycolysis, their ability to produce IFN-γ is markedly compromised. This defect is translational and is regulated by the binding of the glycolysis enzyme GAPDH to AU-rich elements within the 3' UTR of IFN-γ mRNA. GAPDH, by engaging/disengaging glycolysis and through fluctuations in its expression, controls effector cytokine production. Thus, aerobic glycolysis is a metabolically regulated signaling mechanism needed to control cellular function.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Immunometabolism: what is the point of Warburg?Nat Rev Immunol. 2013 Jul;13(7):472. doi: 10.1038/nri3485. Epub 2013 Jun 14. Nat Rev Immunol. 2013. PMID: 23765057 No abstract available.
-
Sweet nothings: sensing of sugar metabolites controls T cell function.Cell Metab. 2013 Jul 2;18(1):7-8. doi: 10.1016/j.cmet.2013.06.009. Cell Metab. 2013. PMID: 23823473 Free PMC article.
-
Immunology. Fueling function over expansion in T cells.Science. 2013 Jul 5;341(6141):37-8. doi: 10.1126/science.1242100. Science. 2013. PMID: 23828930 No abstract available.
References
-
- Anderson P. Post-transcriptional regulons coordinate the initiation and resolution of inflammation. Nat Rev Immunol. 2010;10:24–35. - PubMed
-
- Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439:682–687. - PubMed
-
- Bonafé N, Gilmore-Hebert M, Folk NL, Azodi M, Zhou Y, Chambers SK. Glyceraldehyde-3-phosphate dehydrogenase binds to the AU-Rich 3′ untranslated region of colony-stimulating factor-1 (CSF-1) messenger RNA in human ovarian cancer cells: possible role in CSF-1 posttranscriptional regulation and tumor phenotype. Cancer Res. 2005;65:3762–3771. - PubMed
-
- Bustamente E, Morris HP, Pedersen PL. Hexokinase: the direct link between mitochondrial and glycolytic reactions in rapidly growing cancer cells. Adv Exp Med Biol. 1977;92:363–380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
