

Spatial reorganization of Saccharomyces cerevisiae enolase to alter carbon metabolism under hypoxia
- PMID: 23748432
- PMCID: PMC3754543
- DOI: 10.1128/EC.00093-13
Spatial reorganization of Saccharomyces cerevisiae enolase to alter carbon metabolism under hypoxia
Abstract
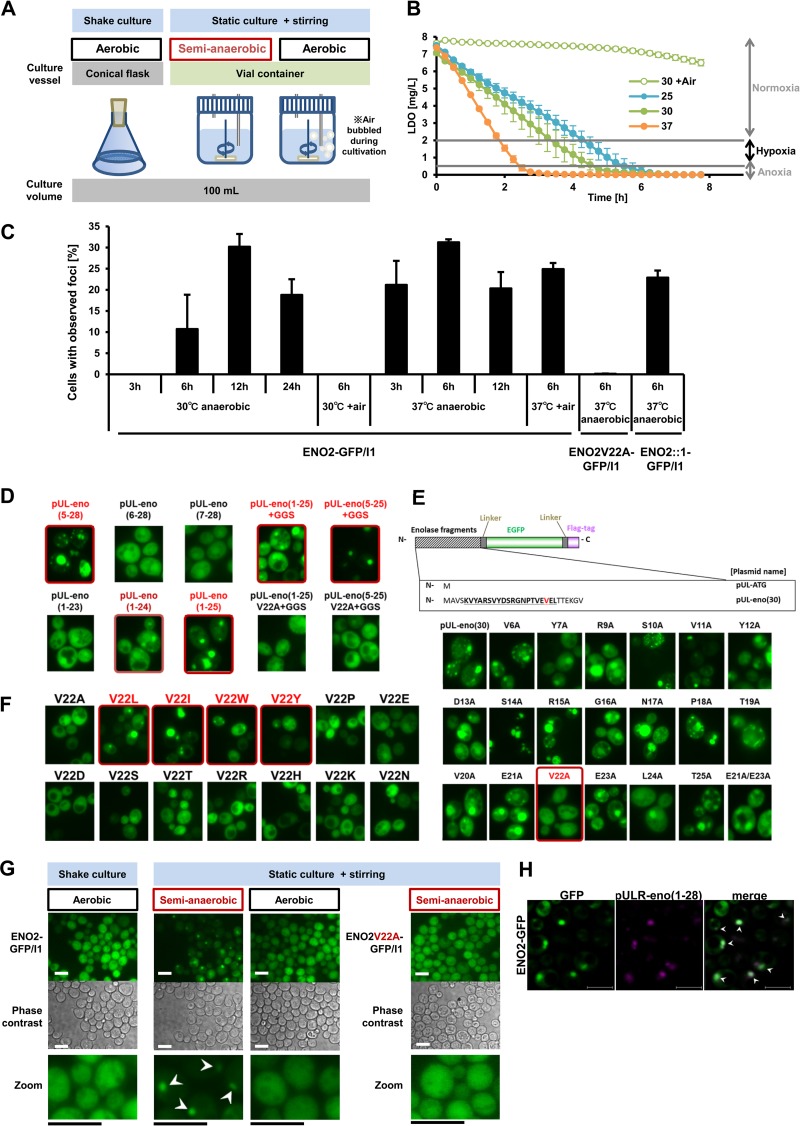
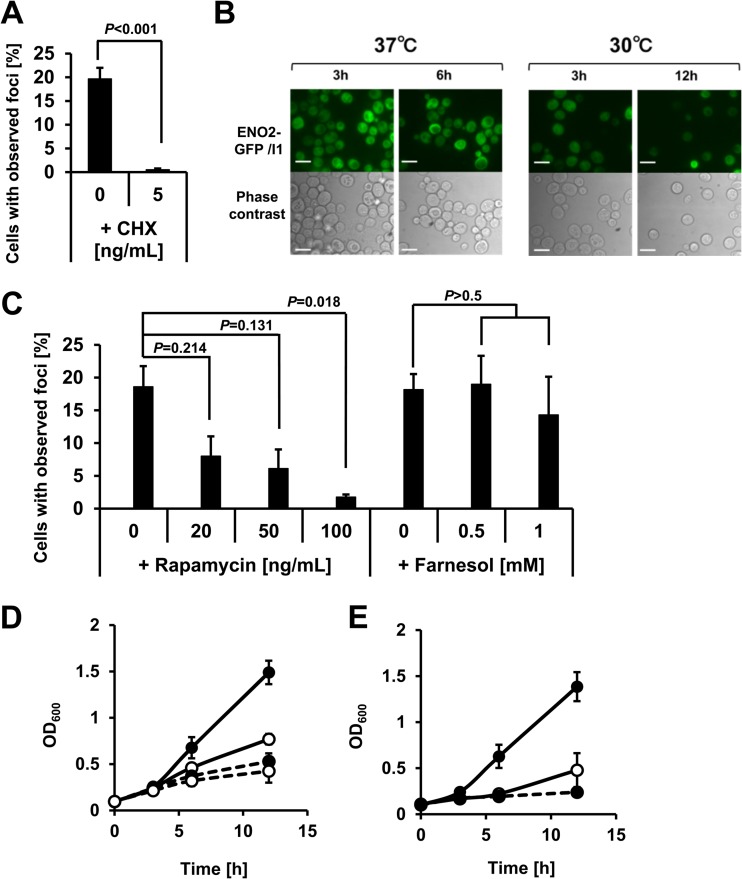
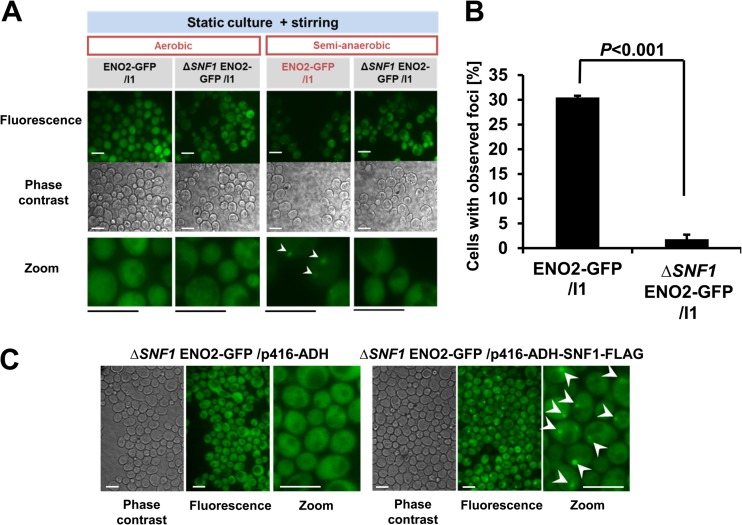
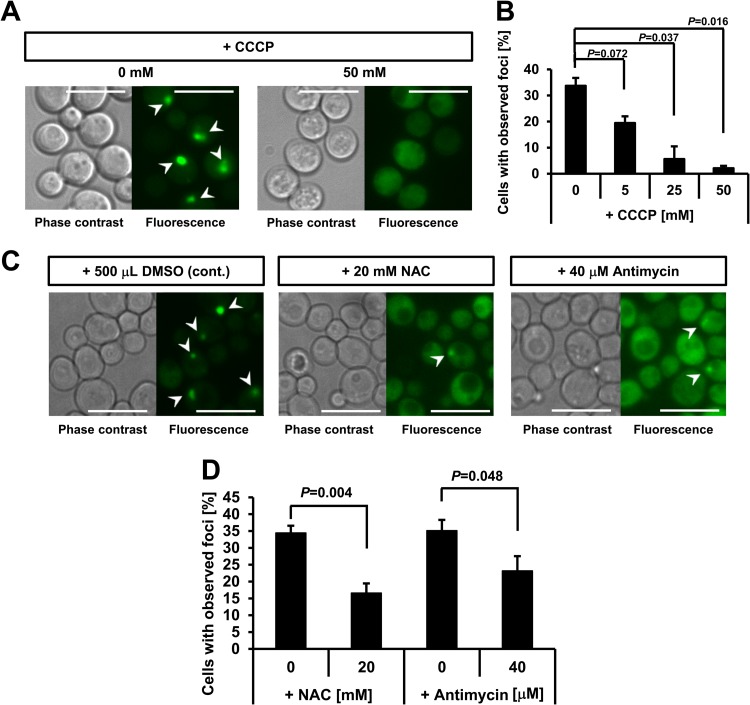
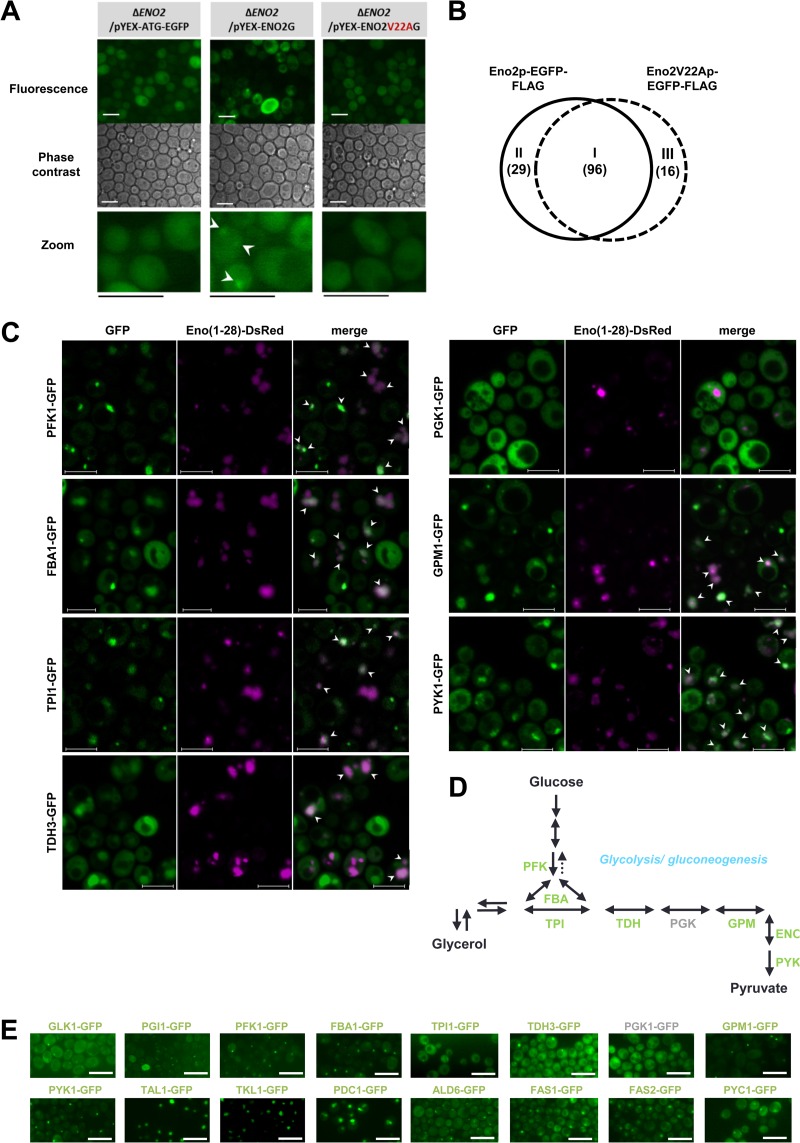
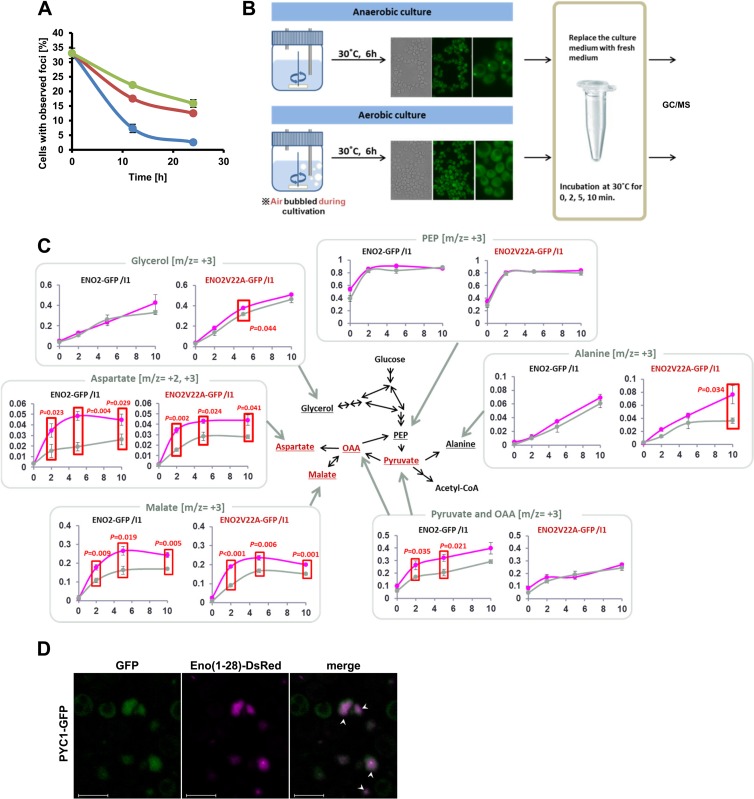
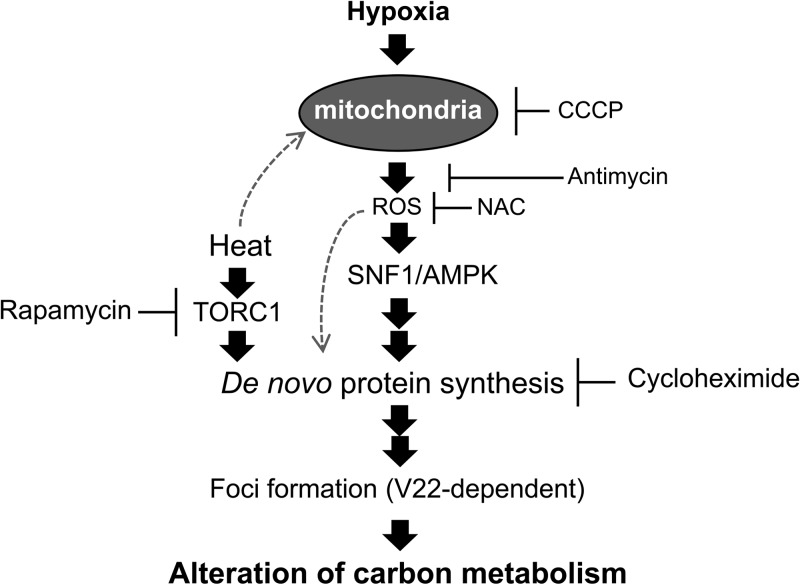
Hypoxia has critical effects on the physiology of organisms. In the yeast Saccharomyces cerevisiae, glycolytic enzymes, including enolase (Eno2p), formed cellular foci under hypoxia. Here, we investigated the regulation and biological functions of these foci. Focus formation by Eno2p was inhibited temperature independently by the addition of cycloheximide or rapamycin or by the single substitution of alanine for the Val22 residue. Using mitochondrial inhibitors and an antioxidant, mitochondrial reactive oxygen species (ROS) production was shown to participate in focus formation. Focus formation was also inhibited temperature dependently by an SNF1 knockout mutation. Interestingly, the foci were observed in the cell even after reoxygenation. The metabolic turnover analysis revealed that [U-(13)C]glucose conversion to pyruvate and oxaloacetate was accelerated in focus-forming cells. These results suggest that under hypoxia, S. cerevisiae cells sense mitochondrial ROS and, by the involvement of SNF1/AMPK, spatially reorganize metabolic enzymes in the cytosol via de novo protein synthesis, which subsequently increases carbon metabolism. The mechanism may be important for yeast cells under hypoxia, to quickly provide both energy and substrates for the biosynthesis of lipids and proteins independently of the tricarboxylic acid (TCA) cycle and also to fit changing environments.
Figures
References
-
- Eby LA, Crowder LB. 2002. Hypoxia-based habitat compression in the Neuse River Estuary: context-dependent shifts in behavioral avoidance thresholds. Can. J. Fish. Aquat. Sci. 59:952–965
-
- Buzzelli CP, Luettich RA, Powers SP, Peterson CH, McNinch JE, Pinckney JL, Paerl HW. 2002. Estimating the spatial extent of bottom-water hypoxia and habitat degradation in a shallow estuary. Mar. Ecol. Prog. Ser. 230:103–112
-
- Hagen T, Taylor CT, Lam F, Moncada S. 2003. Redistribution of intracellular oxygen in hypoxia by nitric oxide: effect on HIF1α. Science 302:1975–1978 - PubMed
-
- Denko NC. 2008. Hypoxia, HIF1 and glucose metabolism in the solid tumour. Nat. Rev. Cancer 8:705–713 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
